

Restoration of adenosine deaminase-deficient human thymocyte development in vitro by inhibition of deoxynucleoside kinases
- PMID: 19018008
- PMCID: PMC2673198
- DOI: 10.4049/jimmunol.181.11.8153
Restoration of adenosine deaminase-deficient human thymocyte development in vitro by inhibition of deoxynucleoside kinases
Abstract
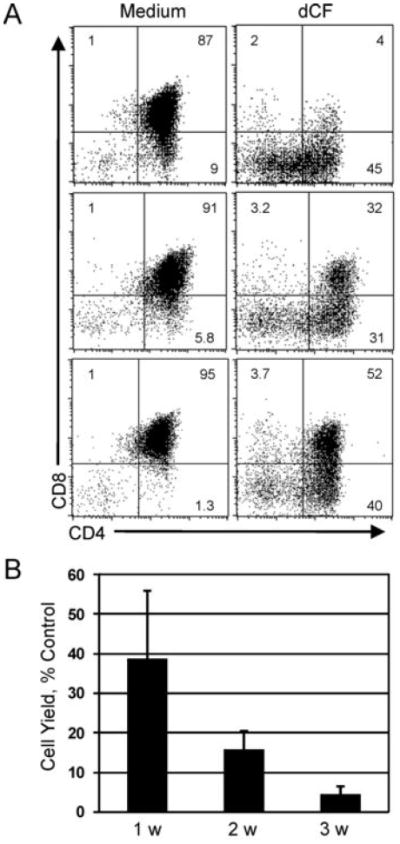
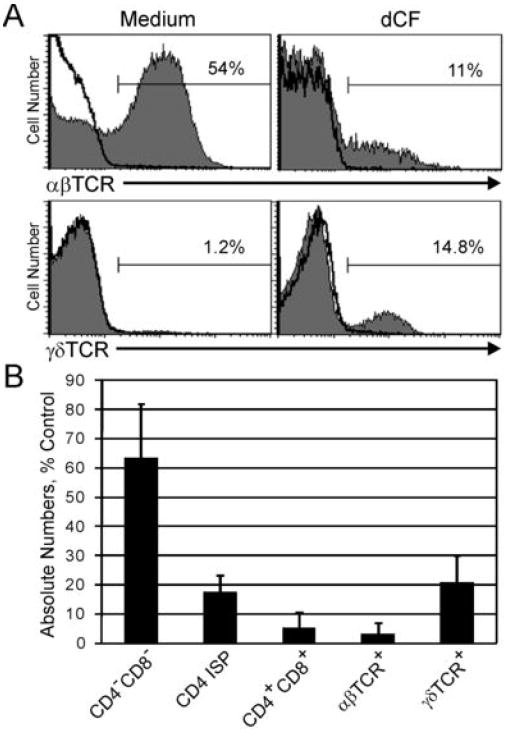
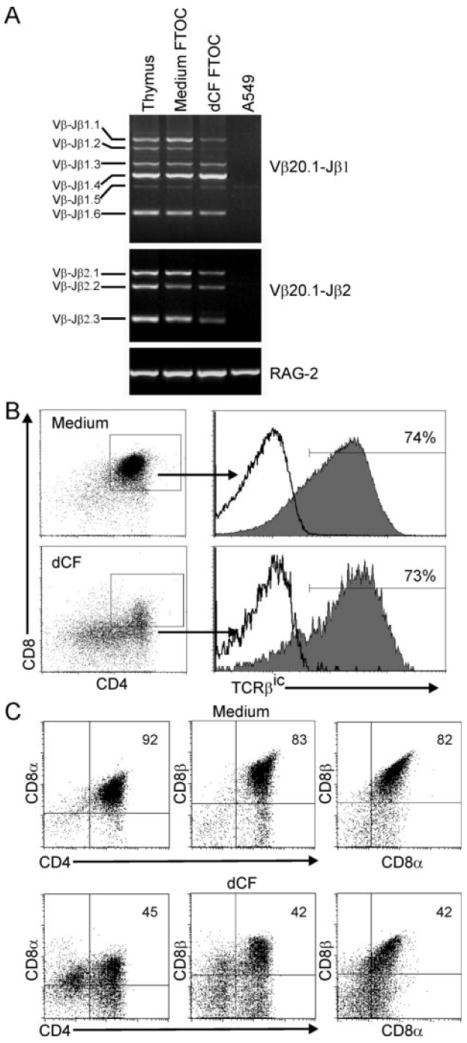
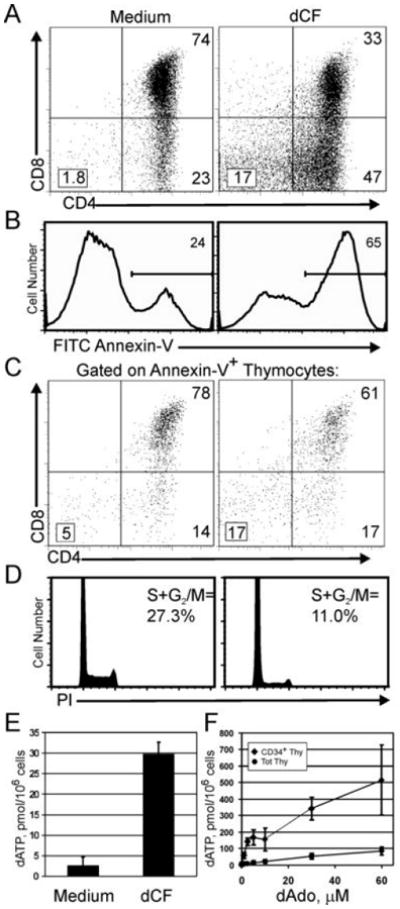
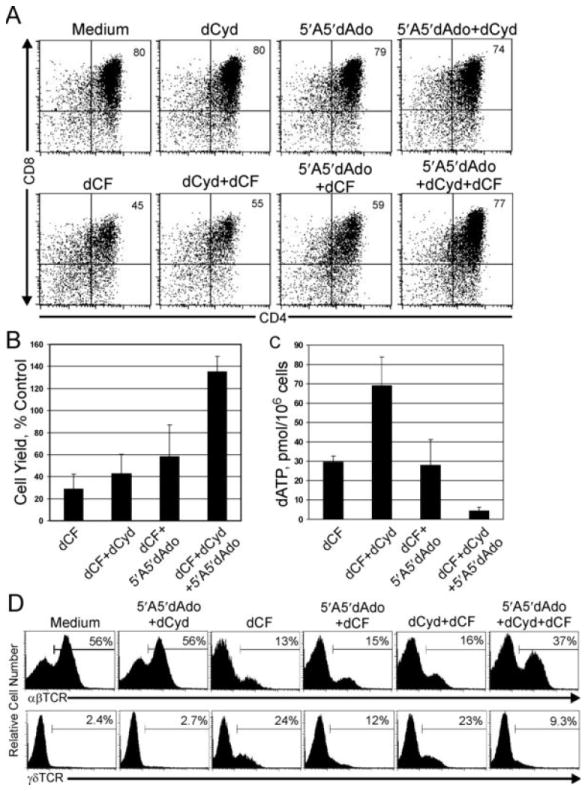
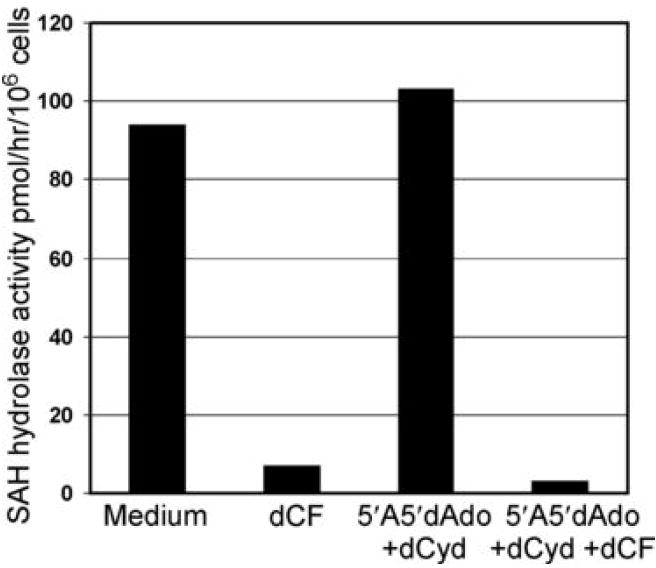
Mutations in the gene encoding adenosine deaminase (ADA), a purine salvage enzyme, lead to immunodeficiency in humans. Although ADA deficiency has been analyzed in cell culture and murine models, information is lacking concerning its impact on the development of human thymocytes. We have used chimeric human/mouse fetal thymic organ culture to study ADA-deficient human thymocyte development in an "in vivo-like" environment where toxic metabolites accumulate in situ. Inhibition of ADA during human thymocyte development resulted in a severe reduction in cellular expansion as well as impaired differentiation, largely affecting mature thymocyte populations. Thymocyte differentiation was not blocked at a discrete stage; rather, the paucity of mature thymocytes was due to the induction of apoptosis as evidenced by activation of caspases and was accompanied by the accumulation of intracellular dATP. Inhibition of adenosine kinase and deoxycytidine kinase prevented the accumulation of dATP and restored thymocyte differentiation and proliferation. Our work reveals that multiple deoxynucleoside kinases are involved in the phosphorylation of deoxyadenosine when ADA is absent, and suggests an alternate therapeutic strategy for treatment of ADA-deficient patients.
Figures
References
-
- Giblett ER, Anderson JE, Cohen F, Pollara B, Meuwissen HJ. Adenosine-deaminase deficiency in two patients with severely impaired cellular immunity. Lancet. 1972;2:1067–1069. - PubMed
-
- Hershfield MS, Mitchell BS. Immunodeficiency diseases caused by adenosine deaminase deficiency and purine nucleoside phosphorylase deficiency. In: Scriver CR, Sly WS, Childs B, Beaudet AL, Valle D, Kinzler K, Vogelstein B, editors. The metabolic and molecular bases of inherited disease. Vol. 2. McGraw-Hill, Inc.; New York: 2001. p. 2585.
-
- Blackburn MR, Kellems RE. Adenosine deaminase deficiency: metabolic basis of immune deficiency and pulmonary inflammation. Adv Immunol. 2005;86:1–41. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
