

A single aromatic residue in transcriptional repressor protein KorA is critical for cooperativity with its co-regulator KorB
- PMID: 19019158
- PMCID: PMC2680271
- DOI: 10.1111/j.1365-2958.2008.06498.x
A single aromatic residue in transcriptional repressor protein KorA is critical for cooperativity with its co-regulator KorB
Abstract
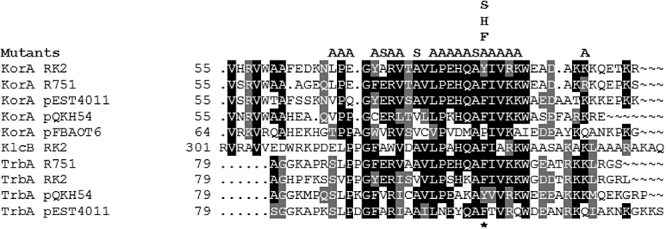
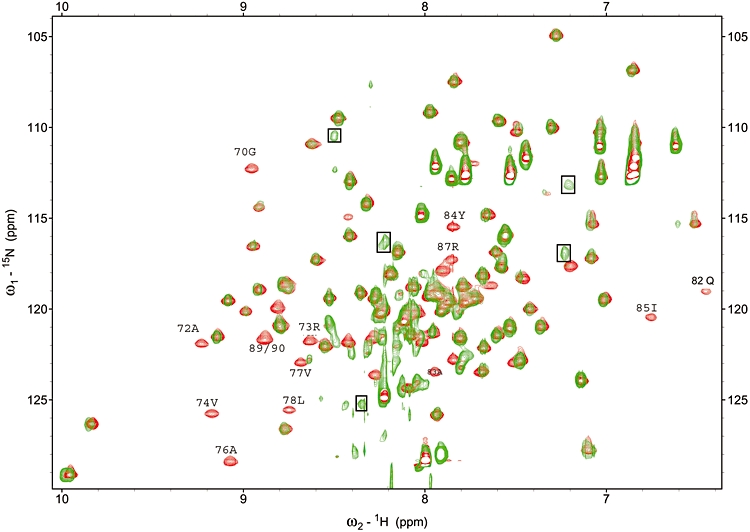
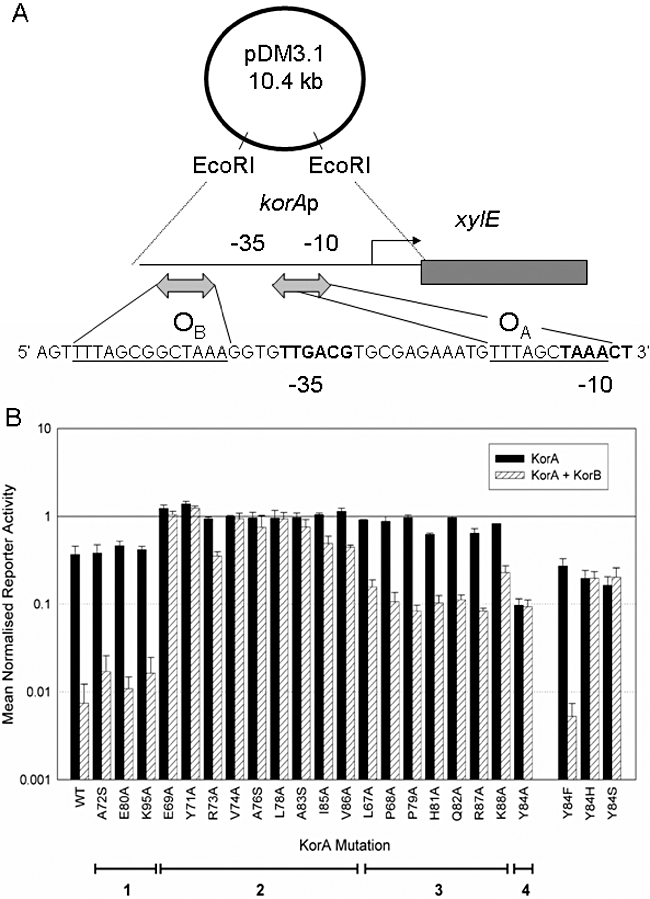
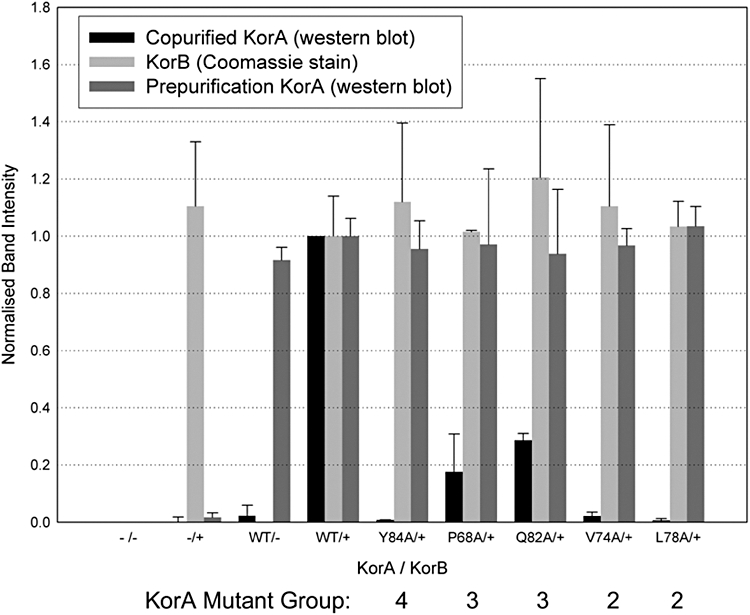
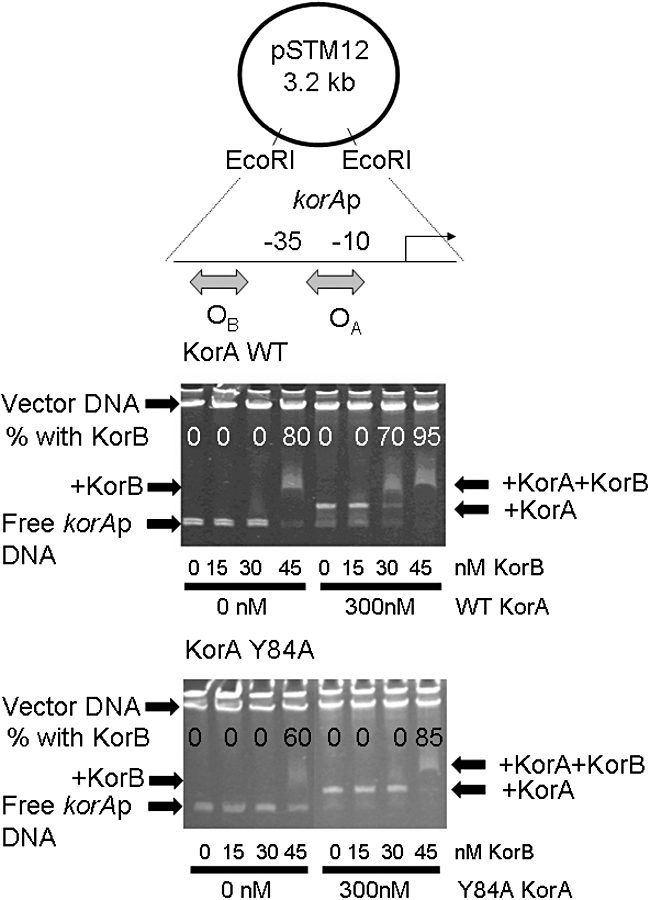
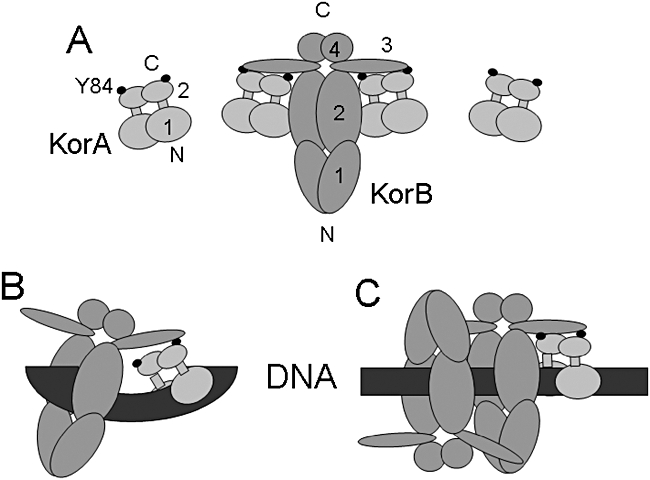
A central feature of broad host range IncP-1 plasmids is the set of regulatory circuits that tightly control plasmid core functions under steady-state conditions. Cooperativity between KorB and either KorA or TrbA repressor proteins is a key element of these circuits and deletion analysis has implicated the conserved C-terminal domain of KorA and TrbA in this interaction. By NMR we show that KorA and KorB interact directly and identify KorA amino acids that are affected on KorB binding. Studies on mutants showed that tyrosine 84 (or phenylalanine, in some alleles) is dispensable for repressor activity but critical for the specific interaction with KorB in both in vivo reporter gene assays and in vitro electrophoretic mobility shift and co-purification assays. This confirms that direct and specific protein-protein interactions are responsible for the cooperativity observed between KorB and its corepressors and lays the basis for determining the biological importance of this cooperativity.
Figures
Similar articles
-
A small protein-protein interaction domain common to KlcB and global regulators KorA and TrbA of promiscuous IncP plasmids.J Mol Biol. 2001 Jun 29;310(1):51-67. doi: 10.1006/jmbi.2001.4729. J Mol Biol. 2001. PMID: 11419936
-
Conserved C-terminal region of global repressor KorA of broad-host-range plasmid RK2 is required for co-operativity between KorA and a second RK2 global regulator, KorB.J Mol Biol. 1999 Jun 4;289(2):211-21. doi: 10.1006/jmbi.1999.2761. J Mol Biol. 1999. PMID: 10366500
-
Flexibility in repression and cooperativity by KorB of broad host range IncP-1 plasmid RK2.J Mol Biol. 2005 Jun 3;349(2):302-16. doi: 10.1016/j.jmb.2005.03.062. Epub 2005 Apr 8. J Mol Biol. 2005. PMID: 15890197
-
Cooperativity between KorB and TrbA repressors of broad-host-range plasmid RK2.J Bacteriol. 2001 Feb;183(3):1022-31. doi: 10.1128/JB.183.3.1022-1031.2001. J Bacteriol. 2001. PMID: 11208801 Free PMC article.
-
Mutational analysis of the global regulator KorA of broad-host-range plasmid RK2.J Mol Biol. 1998 Aug 21;281(3):453-63. doi: 10.1006/jmbi.1998.1956. J Mol Biol. 1998. PMID: 9698561
Cited by
-
Adaptation for protein synthesis efficiency in a naturally occurring self-regulating operon.PLoS One. 2012;7(11):e49678. doi: 10.1371/journal.pone.0049678. Epub 2012 Nov 20. PLoS One. 2012. PMID: 23185406 Free PMC article.
-
Negative feedback and transcriptional overshooting in a regulatory network for horizontal gene transfer.PLoS Genet. 2014 Feb 27;10(2):e1004171. doi: 10.1371/journal.pgen.1004171. eCollection 2014 Feb. PLoS Genet. 2014. PMID: 24586200 Free PMC article.
-
Global Transcriptional Regulation of Backbone Genes in Broad-Host-Range Plasmid RA3 from the IncU Group Involves Segregation Protein KorB (ParB Family).Appl Environ Microbiol. 2016 Apr 4;82(8):2320-2335. doi: 10.1128/AEM.03541-15. Print 2016 Apr. Appl Environ Microbiol. 2016. PMID: 26850301 Free PMC article.
-
Flexibility of KorA, a plasmid-encoded, global transcription regulator, in the presence and the absence of its operator.Nucleic Acids Res. 2016 Jun 2;44(10):4947-56. doi: 10.1093/nar/gkw191. Epub 2016 Mar 25. Nucleic Acids Res. 2016. PMID: 27016739 Free PMC article.
-
Evolution of a Plasmid Regulatory Circuit Ameliorates Plasmid Fitness Cost.bioRxiv [Preprint]. 2024 May 30:2024.02.05.579024. doi: 10.1101/2024.02.05.579024. bioRxiv. 2024. Update in: Mol Biol Evol. 2025 Apr 1;42(4):msaf062. doi: 10.1093/molbev/msaf062. PMID: 38370613 Free PMC article. Updated. Preprint.
References
-
- Adamczyk M, Dolowy P, Jonczyk M, Thomas CM, Jagura-Burdzy G. The kfrA gene is the first in a tricistronic operon required for survival of IncP-1 plasmid R751. Microbiol-SGM. 2006;152:1621–1637. - PubMed
-
- Bailey S, Eliason WK, Steitz TA. Structure of hexameric DnaB helicase and its complex with a domain of DnaG primase. Science. 2007;318:459–463. - PubMed
-
- Barnard A, Wolfe A, Busby S. Regulation at complex bacterial promoters: how bacteria use different promoter organizations to produce different regulatory outcomes. Curr Opin Microbiol. 2004;7:102–108. - PubMed
-
- Batt SM. The active partitioning system of plasmid RK2. PhD Thesis. Birmingham: University of Birmingham.
-
- Bell CE, Frescura P, Hochschild A, Lewis M. Crystal structure of the lambda repressor C-terminal domain provides a model for cooperative operator binding. Cell. 2000;101:801–811. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
