

HLA-associated clinical progression correlates with epitope reversion rates in early human immunodeficiency virus infection
- PMID: 19019964
- PMCID: PMC2620910
- DOI: 10.1128/JVI.01545-08
HLA-associated clinical progression correlates with epitope reversion rates in early human immunodeficiency virus infection
Abstract
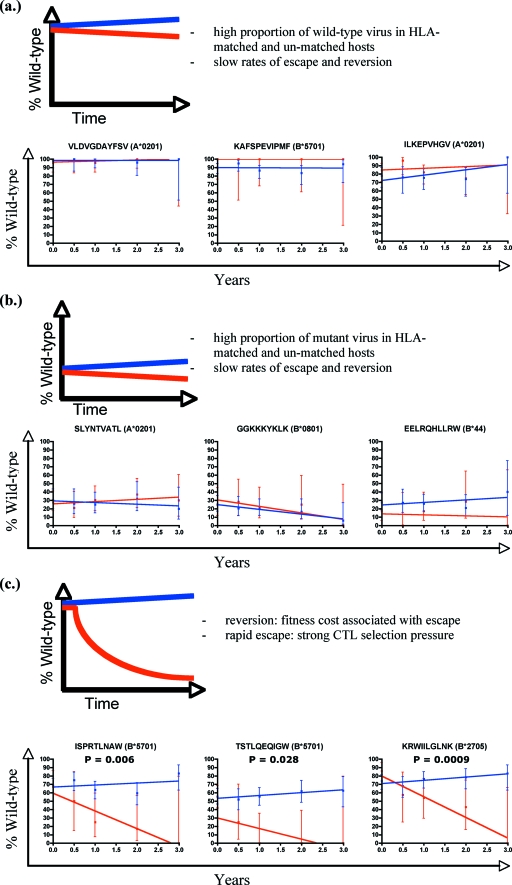
Human immunodeficiency virus type 1 (HIV-1) can evade immunity shortly after transmission to a new host but the clinical significance of this early viral adaptation in HIV infection is not clear. We present an analysis of sequence variation from a longitudinal cohort study of HIV adaptation in 189 acute seroconverters followed for up to 3 years. We measured the rates of variation within well-defined epitopes to determine associations with the HLA-linked hazard of disease progression. We found early reversion across both the gag and pol genes, with a 10-fold faster rate of escape in gag (2.2 versus 0.27 forward mutations/1,000 amino acid sites). For most epitopes (23/34), variation in the HLA-matched and HLA-unmatched controls was similar. For a minority of epitopes (8/34, and generally associated with HLA class I alleles that confer clinical benefit), new variants appeared early and consistently over the first 3 years of infection. Reversion occurred early at a rate which was HLA-dependent and correlated with the HLA class 1-associated relative hazard of disease progression and death (P = 0.0008), reinforcing the association between strong cytotoxic T-lymphocyte responses, viral fitness, and disease status. These data provide a comprehensive overview of viral adaptation in the first 3 years of infection. Our findings of HLA-dependent reversion suggest that costs are borne by some escape variants which may benefit the host, a finding contrary to a simple immune evasion paradigm. These epitopes, which are both strongly and frequently recognized, and for which escape involves a high cost to the virus, have the potential to optimize vaccine design.
Figures




References
-
- Addo, M. M., X. G. Yu, A. Rathod, D. Cohen, R. L. Eldridge, D. Strick, M. N. Johnston, C. Corcoran, A. G. Wurcel, C. A. Fitzpatrick, M. E. Feeney, W. R. Rodriguez, N. Basgoz, R. Draenert, D. R. Stone, C. Brander, P. J. Goulder, E. S. Rosenberg, M. Altfeld, and B. D. Walker. 2003. Comprehensive epitope analysis of human immunodeficiency virus type 1 (HIV-1)-specific T-cell responses directed against the entire expressed HIV-1 genome demonstrate broadly directed responses, but no correlation to viral load. J. Virol. 772081-2092. - PMC - PubMed
-
- Almeida, J. R., D. A. Price, L. Papagno, Z. A. Arkoub, D. Sauce, E. Bornstein, T. E. Asher, A. Samri, A. Schnuriger, I. Theodorou, D. Costagliola, C. Rouzioux, H. Agut, A. G. Marcelin, D. Douek, B. Autran, and V. Appay. 2007. Superior control of HIV-1 replication by CD8+ T cells is reflected by their avidity, polyfunctionality, and clonal turnover. J. Exp. Med. 2042473-2485. - PMC - PubMed
-
- Altfeld, M., E. T. Kalife, Y. Qi, H. Streeck, M. Lichterfeld, M. N. Johnston, N. Burgett, M. E. Swartz, A. Yang, G. Alter, X. G. Yu, A. Meier, J. K. Rockstroh, T. M. Allen, H. Jessen, E. S. Rosenberg, M. Carrington, and B. D. Walker. 2006. HLA alleles associated with delayed progression to AIDS contribute strongly to the initial CD8+ T cell response against HIV-1. PLoS Med. 3e403. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
