

Specific glial populations regulate hippocampal morphogenesis
- PMID: 19020026
- PMCID: PMC6671696
- DOI: 10.1523/JNEUROSCI.4000-08.2008
Specific glial populations regulate hippocampal morphogenesis
Abstract
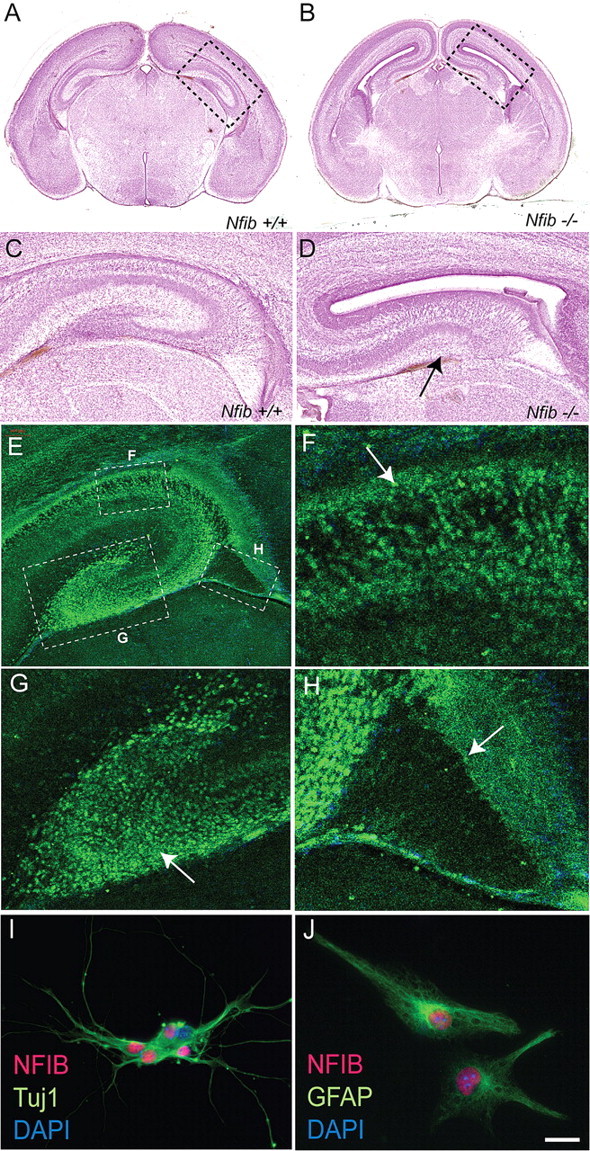
The hippocampus plays an integral role in spatial navigation, learning and memory, and is a major site for adult neurogenesis. Critical to these functions is the proper organization of the hippocampus during development. Radial glia are known to regulate hippocampal formation, but their precise function in this process is yet to be defined. We find that in Nuclear Factor I b (Nfib)-deficient mice, a subpopulation of glia from the ammonic neuroepithelium of the hippocampus fail to develop. This results in severe morphological defects, including a failure of the hippocampal fissure, and subsequently the dentate gyrus, to form. As in wild-type mice, immature nestin-positive glia, which encompass all types of radial glia, populate the hippocampus in Nfib-deficient mice at embryonic day 15. However, these fail to mature into GLAST- and GFAP-positive glia, and the supragranular glial bundle is absent. In contrast, the fimbrial glial bundle forms, but alone is insufficient for proper hippocampal morphogenesis. Dentate granule neurons are present in the mutant hippocampus but their migration is aberrant, likely resulting from the lack of the complete radial glial scaffold usually provided by both glial bundles. These data demonstrate a role for Nfib in hippocampal fissure and dentate gyrus formation, and that distinct glial bundles are critical for correct hippocampal morphogenesis.
Figures









References
-
- Altman J, Bayer SA. Migration and distribution of two populations of hippocampal granule cell precursors during the perinatal and postnatal periods. J Comp Neurol. 1990a;301:365–381. - PubMed
-
- Altman J, Bayer SA. Mosaic organization of the hippocampal neuroepithelium and the multiple germinal sources of dentate granule cells. J Comp Neurol. 1990b;301:325–342. - PubMed
-
- Bettler B, Boulter J, Hermans-Borgmeyer I, O'Shea-Greenfield A, Deneris ES, Moll C, Borgmeyer U, Hollmann M, Heinemann S. Cloning of a novel glutamate receptor subunit, GluR5: expression in the nervous system during development. Neuron. 1990;5:583–595. - PubMed
-
- Bielle F, Griveau A, Narboux-Nême N, Vigneau S, Sigrist M, Arber S, Wassef M, Pierani A. Multiple origins of Cajal-Retzius cells at the borders of the developing pallium. Nat Neurosci. 2005;8:1002–1012. - PubMed
-
- Bulchand S, Grove EA, Porter FD, Tole S. LIM-homeodomain gene Lhx2 regulates the formation of the cortical hem. Mech Dev. 2001;100:165–175. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous