

Eight genes are required for functional reconstitution of the Caenorhabditis elegans levamisole-sensitive acetylcholine receptor
- PMID: 19020092
- PMCID: PMC2587545
- DOI: 10.1073/pnas.0806933105
Eight genes are required for functional reconstitution of the Caenorhabditis elegans levamisole-sensitive acetylcholine receptor
Abstract
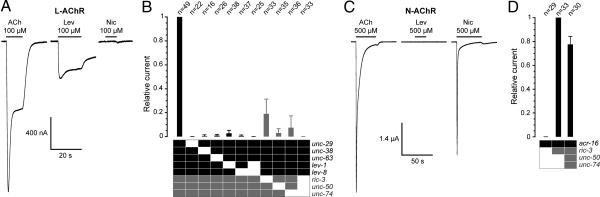
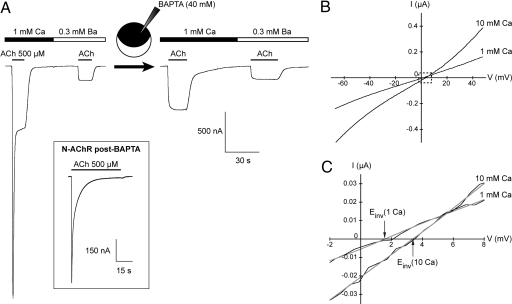
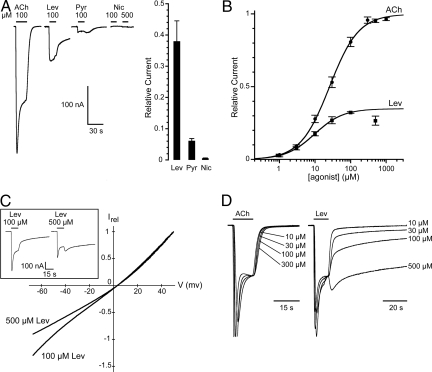
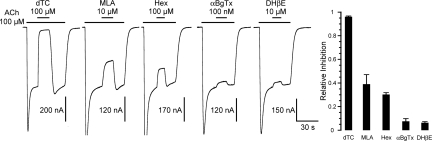
Levamisole-sensitive acetylcholine receptors (L-AChRs) are ligand-gated ion channels that mediate excitatory neurotransmission at the neuromuscular junctions of nematodes. They constitute a major drug target for anthelminthic treatments because they can be activated by nematode-specific cholinergic agonists such as levamisole. Genetic screens conducted in Caenorhabditis elegans for resistance to levamisole toxicity identified genes that are indispensable for the biosynthesis of L-AChRs. These include 5 genes encoding distinct AChR subunits and 3 genes coding for ancillary proteins involved in assembly and trafficking of the receptors. Despite extensive analysis of L-AChRs in vivo, pharmacological and biophysical characterization of these receptors has been greatly hampered by the absence of a heterologous expression system. Using Xenopus laevis oocytes, we were able to reconstitute functional L-AChRs by coexpressing the 5 distinct receptor subunits and the 3 ancillary proteins. Strikingly, this system recapitulates the genetic requirements for receptor expression in vivo because omission of any of these 8 genes dramatically impairs L-AChR expression. We demonstrate that 3 alpha- and 2 non-alpha-subunits assemble into the same receptor. Pharmacological analysis reveals that the prototypical cholinergic agonist nicotine is unable to activate L-AChRs but rather acts as a potent allosteric inhibitor. These results emphasize the role of ancillary proteins for efficient expression of recombinant neurotransmitter receptors and open the way for in vitro screening of novel anthelminthic agents.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Bethony J, et al. Soil-transmitted helminth infections: Ascariasis, trichuriasis, and hookworm. Lancet. 2006;367:1521–1532. - PubMed
-
- Keiser J, Utzinger J. Efficacy of current drugs against soil-transmitted helminth infections: Systematic review and meta-analysis. J Am Med Assoc. 2008;299:1937–1948. - PubMed
-
- Rayes D, De Rosa MJ, Bartos M, Bouzat C. Molecular basis of the differential sensitivity of nematode and mammalian muscle to the anthelmintic agent levamisole. J Biol Chem. 2004;279:36372–36381. - PubMed
-
- Jones AK, Sattelle DB. Functional genomics of the nicotinic acetylcholine receptor gene family of the nematode, Caenorhabditis elegans. Bioessays. 2004;26:39–49. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
