

Control of WHITE COLLAR localization by phosphorylation is a critical step in the circadian negative feedback process
- PMID: 19020516
- PMCID: PMC2609740
- DOI: 10.1038/emboj.2008.245
Control of WHITE COLLAR localization by phosphorylation is a critical step in the circadian negative feedback process
Abstract
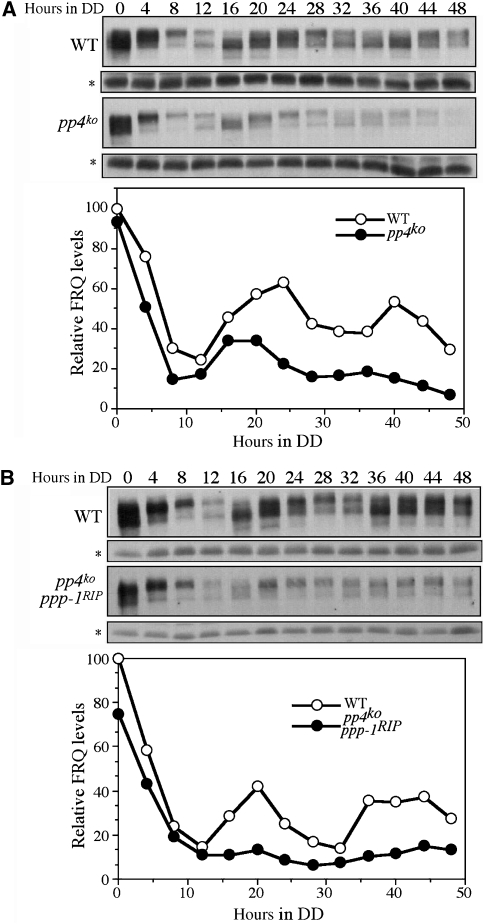
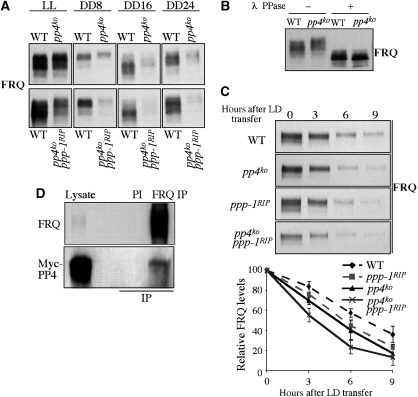
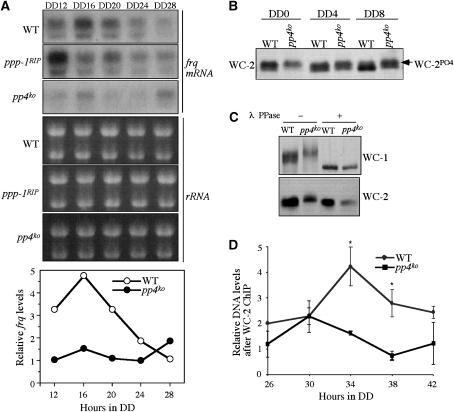
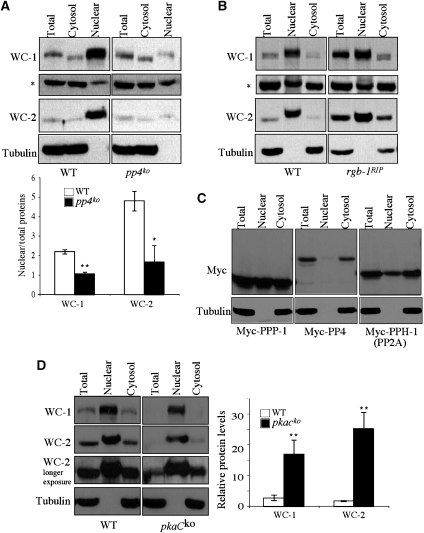
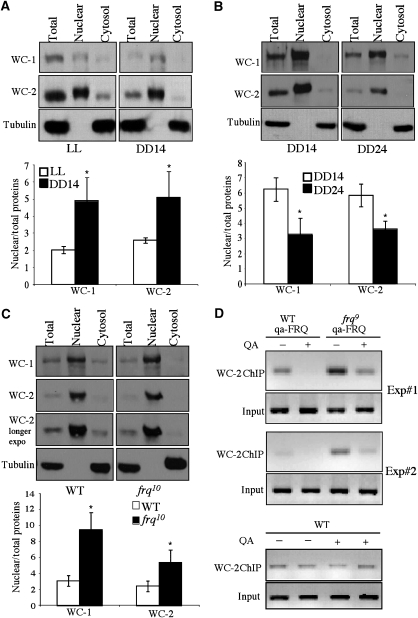
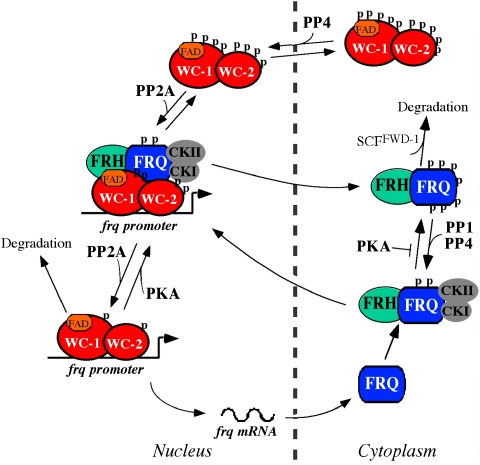
Reversible protein phosphorylation has critical functions in the eukaryotic circadian negative feedback loops. In Neurospora, the FREQUENCY protein closes the circadian negative feedback loop by promoting the phosphorylation of its transcription activator, the WHITE COLLAR complex (WCC) and consequently inhibiting WCC activity. Here we show that protein phosphatase 4 is a novel component of the Neurospora clock by regulating both processes of the circadian negative feedback loop. The disruption of pp4 results in short period rhythms with low amplitude. In addition to its role in regulating FRQ phosphorylation and stability, PP4 also dephosphorylates and activates WCC. In contrast to PP2A, another phosphatase that activates WCC, PP4 has a major function in promoting nuclear entry of WCC. PKA, a WC kinase, inhibits WC nuclear localization. Furthermore, the FRQ-dependent WC phosphorylation promotes WCC cytosolic localization. Together, these results revealed WCC nucleocytoplasmic shuttling as an important step in the circadian negative feedback process and delineated the FRQ-dependent WCC inhibition as a two-step process: the inhibition of WCC DNA-binding activity followed by sequestration of WCC into the cytoplasm.
Figures

References
-
- Allada R, Emery P, Takahashi JS, Rosbash M (2001) Stopping time: the genetics of fly and mouse circadian clocks. Annu Rev Neurosci 24: 1091–1119 - PubMed
-
- Aronson B, Johnson K, Loros JJ, Dunlap JC (1994a) Negative feedback defining a circadian clock: autoregulation in the clock gene frequency. Science 263: 1578–1584 - PubMed
-
- Belden WJ, Loros JJ, Dunlap JC (2007) Execution of the circadian negative feedback loop in Neurospora requires the ATP-dependent chromatin-remodeling enzyme CLOCKSWITCH. Mol Cell 25: 587–600 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
