

Mouse Leydig cells express multiple P2X receptor subunits
- PMID: 19020992
- PMCID: PMC2717309
- DOI: 10.1007/s11302-008-9128-9
Mouse Leydig cells express multiple P2X receptor subunits
Abstract
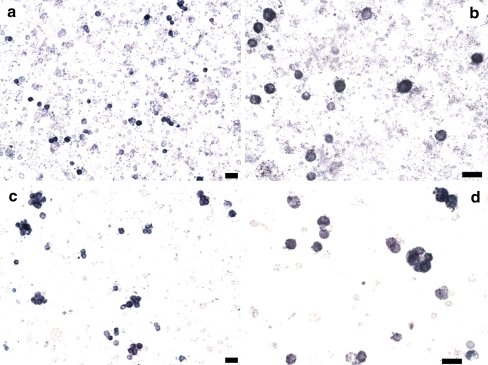
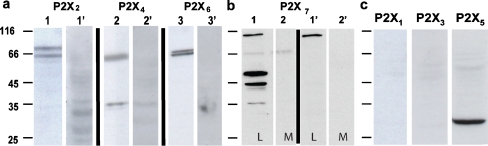
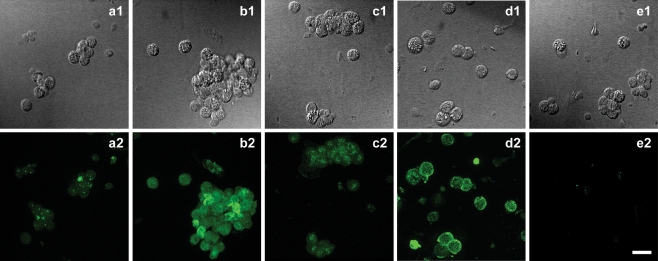
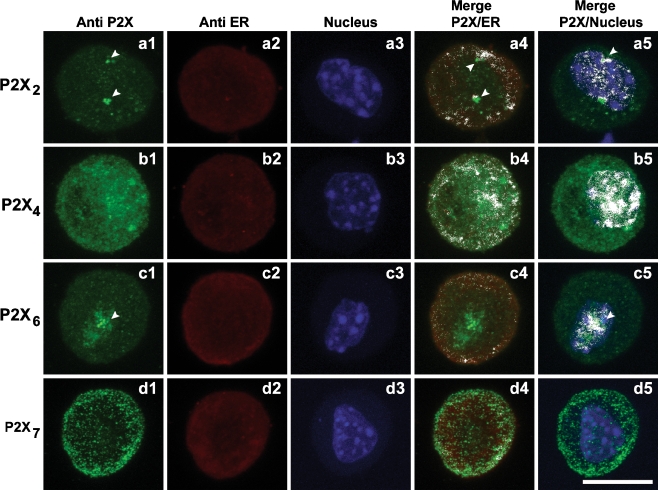
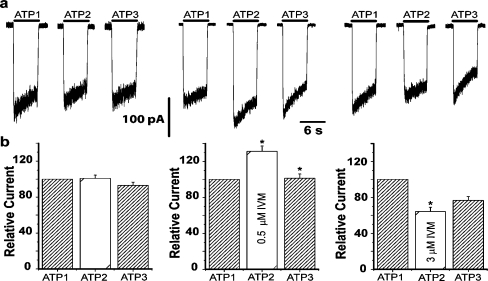
ATP acts on cellular membranes by interacting with P2X (ionotropic) and P2Y (metabotropic) receptors. Seven homomeric P2X receptors (P2X(1)-P2X(7)) and seven heteromeric receptors (P2X(1/2), P2X(1/4), P2X(1/5), P2X(2/3), P2X(2/6), P2X(4/6), P2X(4/7)) have been described. ATP treatment of Leydig cells leads to an increase in [Ca(2+)](i) and testosterone secretion, supporting the hypothesis that Ca(2+) signaling through purinergic receptors contributes to the process of testosterone secretion in these cells. Mouse Leydig cells have P2X receptors with a pharmacological and biophysical profile resembling P2X(2). In this work, we describe the presence of several P2X receptor subunits in mouse Leydig cells. Western blot experiments showed the presence of P2X(2), P2X(4), P2X(6), and P2X(7) subunits. These results were confirmed by immunofluorescence. Functional results support the hypothesis that heteromeric receptors are present in these cells since 0.5 muM ivermectin induced an increase (131.2 +/- 5.9%) and 3 muM ivermectin a decrease (64.2 +/- 4.8%) in the whole-cell currents evoked by ATP. These results indicate the presence of functional P2X(4) subunits. P2X(7) receptors were also present, but they were non-functional under the present conditions because dye uptake experiments with Lucifer yellow and ethidium bromide were negative. We conclude that a heteromeric channel, possibly P2X(2/4/6), is present in Leydig cells, but with an electrophysiological and pharmacological phenotype characteristic of the P2X(2) subunit.
Figures

References
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1016/j.jmb.2004.06.092', 'is_inner': False, 'url': 'https://doi.org/10.1016/j.jmb.2004.06.092'}, {'type': 'PubMed', 'value': '15313628', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/15313628/'}]}
- Aschrafi A, Sadtler S, Niculescu C, Rettinger J, Schmalzing G (2004) Trimeric architecture of homomeric P2X2 and heteromeric P2X1+2 receptor subtypes. J Mol Biol 342:333–343 doi:10.1016/j.jmb.2004.06.092 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1016/0014-5793(95)01203-Q', 'is_inner': False, 'url': 'https://doi.org/10.1016/0014-5793(95)01203-q'}, {'type': 'PubMed', 'value': '7498461', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/7498461/'}]}
- Bo X, Zhang Y, Nassar M, Burnstock G, Schoepfer R (1995) A P2X purinoceptor cDNA conferring a novel pharmacological profile. FEBS Lett 375:129–133 doi:10.1016/0014-5793(95)01203-Q - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1038/371519a0', 'is_inner': False, 'url': 'https://doi.org/10.1038/371519a0'}, {'type': 'PubMed', 'value': '7523952', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/7523952/'}]}
- Brake AJ, Wagenbach MJ, Julius D (1994) New structural motif for ligand-gated ion channel defined by ionotropic ATP receptor. Nature 371:519–523 doi:10.1038/371519a0 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1016/j.imlet.2007.07.015', 'is_inner': False, 'url': 'https://doi.org/10.1016/j.imlet.2007.07.015'}, {'type': 'PubMed', 'value': '17825926', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/17825926/'}]}
- Brône B, Moechars D, Marrannes R, Mercken M, Meert T (2007) P2X currents in peritoneal macrophages of wild type and P2X4 −/− mice. Immunol Lett 113:83–89 doi:10.1016/j.imlet.2007.07.015 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1124/jpet.300.2.673', 'is_inner': False, 'url': 'https://doi.org/10.1124/jpet.300.2.673'}, {'type': 'PMC', 'value': 'PMC5577565', 'is_inner': False, 'url': 'https://pmc.ncbi.nlm.nih.gov/articles/PMC5577565/'}, {'type': 'PubMed', 'value': '11805232', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/11805232/'}]}
- Brown SG, Townsend-nicholson A, Jacobson KA, Burnstock G, King BF (2002) Heteromultimeric P2X1/2 receptors show a novel sensitivity to extracellular pH. J Pharmacol Exp Ther 300:673–680 doi:10.1124/jpet.300.2.673 - PMC - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
