

doi: 10.1111/j.1742-4658.2008.06693.x.
A decade of Cdc14--a personal perspective. Delivered on 9 July 2007 at the 32nd FEBS Congress in Vienna, Austria
Affiliations
- PMID: 19021755
- PMCID: PMC2742780
- DOI: 10.1111/j.1742-4658.2008.06693.x
Item in Clipboard
A decade of Cdc14--a personal perspective. Delivered on 9 July 2007 at the 32nd FEBS Congress in Vienna, Austria
FEBS J.
2008 Dec.
Abstract
In budding yeast, the protein phosphatase Cdc14 is a key regulator of late mitotic events. Research over the last decade has revealed many of its functions and today we know that this protein phosphatase orchestrates several aspects of chromosome segregation and is the key trigger of exit from mitosis. Elucidation of the mechanisms controlling Cdc14 activity through nucleolar sequestration now serves as a paradigm for how regulation of the subcellular localization of proteins regulates protein function. Here I review these findings focusing on how discoveries in my laboratory helped elucidate the function and regulation of Cdc14.
Figures
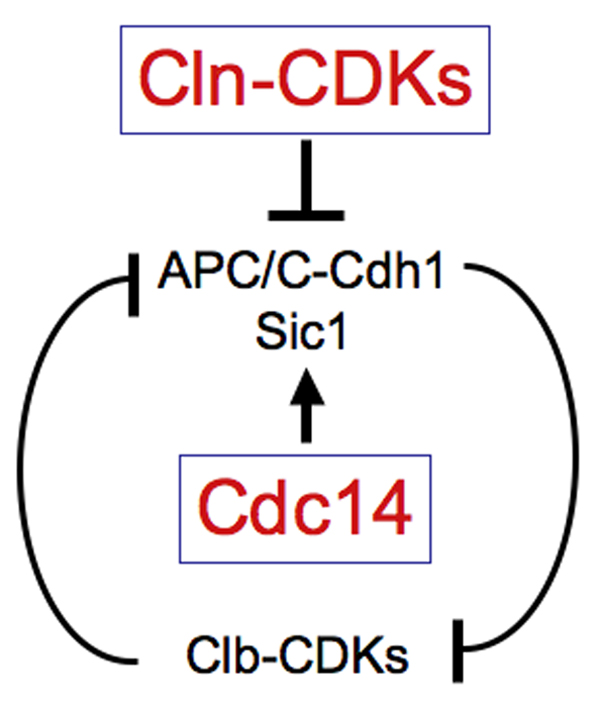
Clb-CDKs and the factors that inactivate it mutually inhibit each other. During S phase and early mitosis, Clb-CDKs inhibit APC/C-Cdh1 by preventing the association of Cdh1 with APC/C. Clb-CDKs also prevent the accumulation of Sic1. They prevent entry of the transcription factor Swi5 into the nucleus thereby inhibiting SIC1 transcription. They also phopshorylate Sic1, which targets it for degradation. Activation of Cdc14 during anaphase dephosphorylates Cdh1, which causes activation of the APC-Cdh1 and hence mitotic cyclin degradation. Cdc14 also dephosphorylates Swi5 and Sic1, causing SIC1 transcription and Sic1 stabilization, respectively. This leads to the Clb-CDK inhibitors to gain the upper hand, keeping Clb-CDKs in the inactive state. Cln-CDKs break this inhibition once they are active at the G1 – S phase transition. They target Sic1 for degradation and phosphorylate Cdh1, allowing Clb-CDKs to accumulate again.
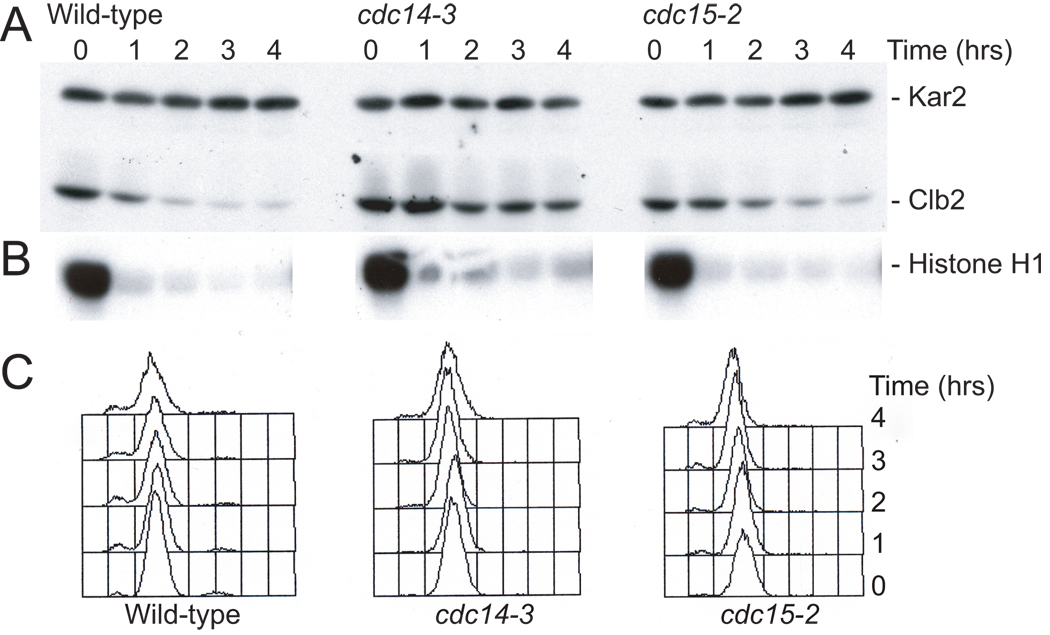
GAL-SIC1 (A810), cdc14-3 GAL-SIC1 (A831) and cdc15-2 GAL-SIC1 (A844) cells were arrested with nocodazole (15µg/ml) for 165 minutes in YEP medium containing 2% raffinose at 23°C. Then the 0 time point was taken and cells were shifted to 37°C for 30 minutes. Then 2% galactose and α–factor (5 µg/ml) were added to induce Sic1 production and to inhibit Cln-CDKs, respectively. 5µg/ml nocodazole was readded at the same time to ensure that microtubules remain depolymerized. Samples were taken at the indicated times after temperature shift to determine the amount of Clb2 protein (A), Clb2-associated histone H1 kinase activity (B) and DNA content (C). Kar2 was used as a loading control in western blots. Methods were as described in Amon (1997).

The degradation of Pds1 and hence activation of Esp1 marks the onset of anaphase. Esp1 then not only cleaves cohesins to bring about sister chromatid segregation but together with Slk19 also promotes the down-regulation of the protein phosphatase PP2A. This allows Clb-CDKs to phosphorylate Cfi1/Net1 and Spo12, which brings about the dissociation of Cdc14 from its inhibitor. Phosphorylation of Cfi1/Net1 directly disrupts the Cdc14 – Cfi1/Net1 complex. Phosphorylation of Spo12 promotes the protein to inhibit Fob1, which inhibits Cdc14 – Cfi1/Net1 dissociation. Movement of the MEN-bearing SPB into the bud, where Lte1 is located, down-regulation of Bub2-Bfa1 activity by Cdc5, inactivation of Kin4 and other unknown signals promote MEN activation during anaphase. Tem1, presumably in its GTP bound form, then activates Cdc15, which activates Dbf2 in a Mob1-dependent manner. Dbf2-Mob1 promotes Cdc14 release from the nucleolus by an unknown mechanism.
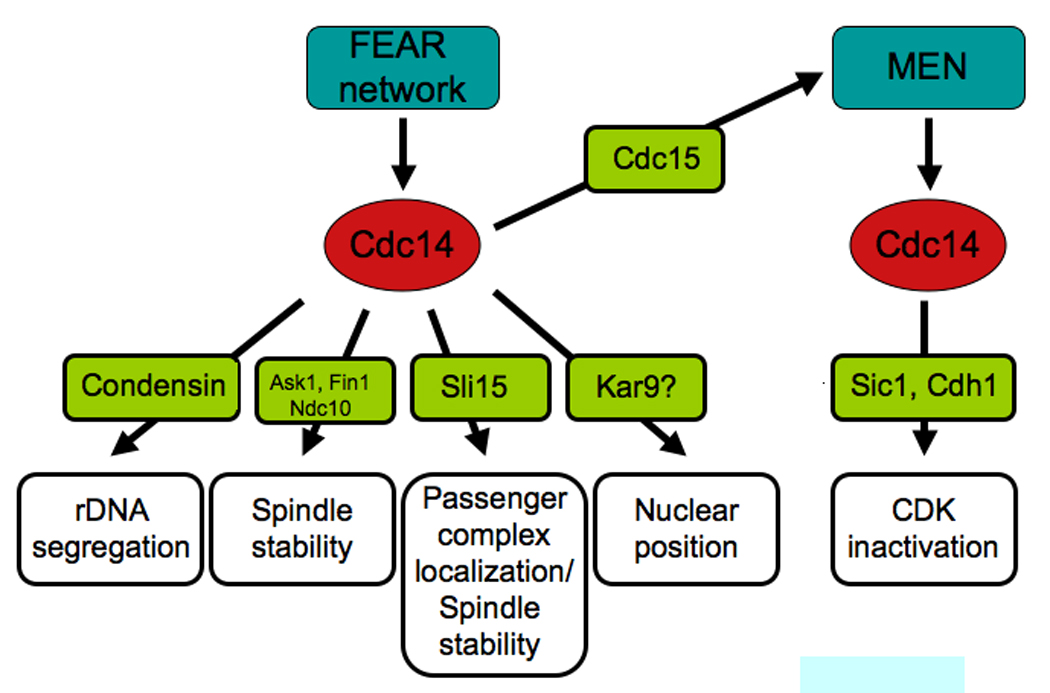
At anaphase onset, Cdc14 is activated by the FEAR network and controls many aspects of anaphase chromosome movement. The protein phosphatase promotes rDNA segregation by targeting condensins to the rDNA. It stabilizes the anaphase spindle by dephosphorylating kinetochore and spindle proteins such as Ask1, Ndc10, Fin1 and the chromosomal passenger complex. Cdc14 promotes the localization of the chromosomal passenger protein complex to the spindle midzone and controls nuclear position. Cdc14 also promotes MEN activity, which is necessary to maintain Cdc14 in the released active state during late stages of anaphase. Once activated the MEN further promotes Cdc14 activity. This sustained activation of Cdc14 then brings about exit from mitosis.

A certain amount of Clb-CDK activity is required for cells to enter mitosis (Threshold 1). The amount of Clb-CDK activity needed for entry into mitosis is not as high as that for anaphase initiation (Threshold 2). This is illustrated by the fact that inactivation of Clb1 – 4 leads to a G2 arrest. Inactivation of Clb2 and Clb1 or Clb2 and Clb3 results in a metaphase delay. The need for increasing amounts of Clb-CDK activity could help establish an order of events as cells progress from G2 into metaphase. Increasing levels of Cdc14 activity and hence decreasing levels of Clb-CDK activity could also help establish order to the progression from metaphase to G1. Once cells enter anaphase Clb-CDK activity needs to decline some for accurate anaphase chromosome movement (Threshold 3). Cdc14 released by the FEAR network brings about this event. For cells to exit from mitosis, Clb-CDK activity needs to be lowered even further (Threshold 4). Eventually all Clb-CDKs are inactivated resetting the cell cycle to a GAP phase state.
References
-
- Bloom J, Cross FR. Multiple levels of cyclin specificity in cell-cycle control. Nat Rev Mol Cell Bio. 2007;8(2):149–160. Review. - PubMed
-
- Murray AW, Solomon MJ, Kirschner MW. The role of cyclin synthesis and degradation in the control of maturation promoting factor activity. Nature. 1989;339(6222):280–286. - PubMed
-
- Irniger S, Piatti S, Michaelis C, Nasmyth K. Genes involved in sister chromatid separation are needed for B-type cyclin proteolysis in budding yeast. Cell. 1995;81(2):269–278. Erratum in: Cell 93(3), 487. - PubMed
-
- King RW, Peters JM, Tugendreich S, Rolfe M, Hieter P, Kirschner MW. A 20S complex containing CDC27 and CDC16 catalyzes the mitosis-specific conjugation of ubiquitin to cyclin B. Cell. 1995;81(2):279–288. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
