

Structure and function of HIV-1 reverse transcriptase: molecular mechanisms of polymerization and inhibition
- PMID: 19022262
- PMCID: PMC2881421
- DOI: 10.1016/j.jmb.2008.10.071
Structure and function of HIV-1 reverse transcriptase: molecular mechanisms of polymerization and inhibition
Abstract
The rapid replication of HIV-1 and the errors made during viral replication cause the virus to evolve rapidly in patients, making the problems of vaccine development and drug therapy particularly challenging. In the absence of an effective vaccine, drugs are the only useful treatment. Anti-HIV drugs work; so far drug therapy has saved more than three million years of life. Unfortunately, HIV-1 develops resistance to all of the available drugs. Although a number of useful anti-HIV drugs have been approved for use in patients, the problems associated with drug toxicity and the development of resistance means that the search for new drugs is an ongoing process. The three viral enzymes, reverse transcriptase (RT), integrase (IN), and protease (PR) are all good drug targets. Two distinct types of RT inhibitors, both of which block the polymerase activity of RT, have been approved to treat HIV-1 infections, nucleoside analogs (NRTIs) and nonnucleosides (NNRTIs), and there are promising leads for compounds that either block the RNase H activity or block the polymerase in other ways. A better understanding of the structure and function(s) of RT and of the mechanism(s) of inhibition can be used to generate better drugs; in particular, drugs that are effective against the current drug-resistant strains of HIV-1.
Figures
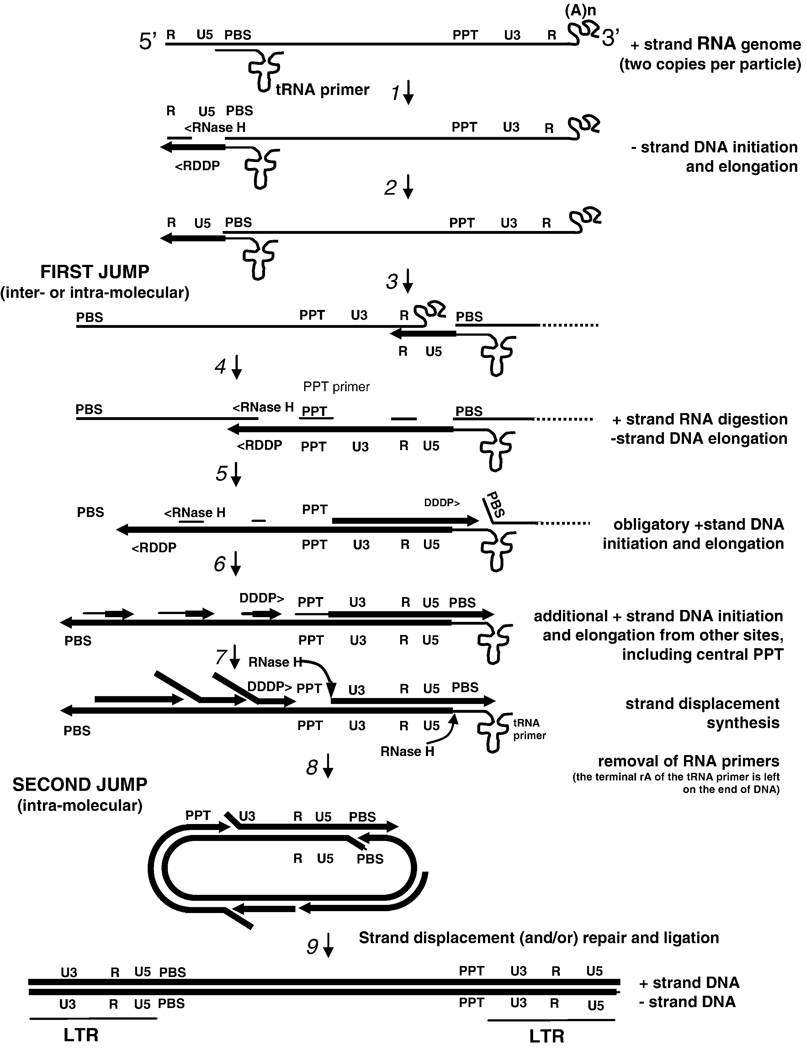
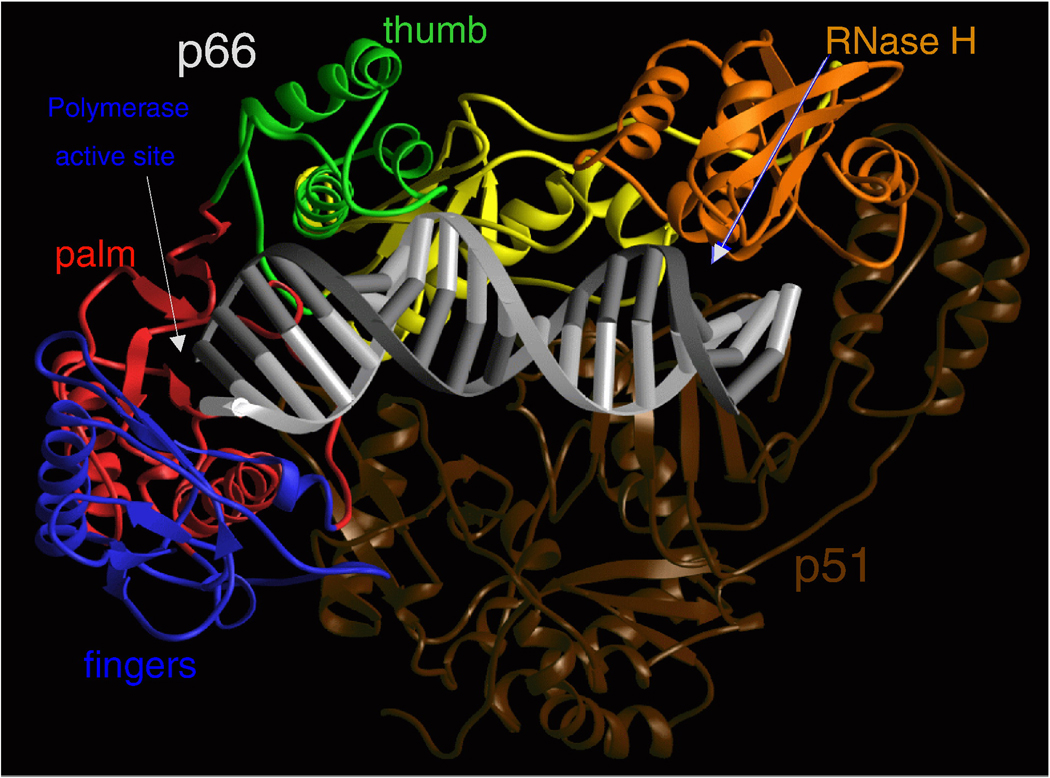
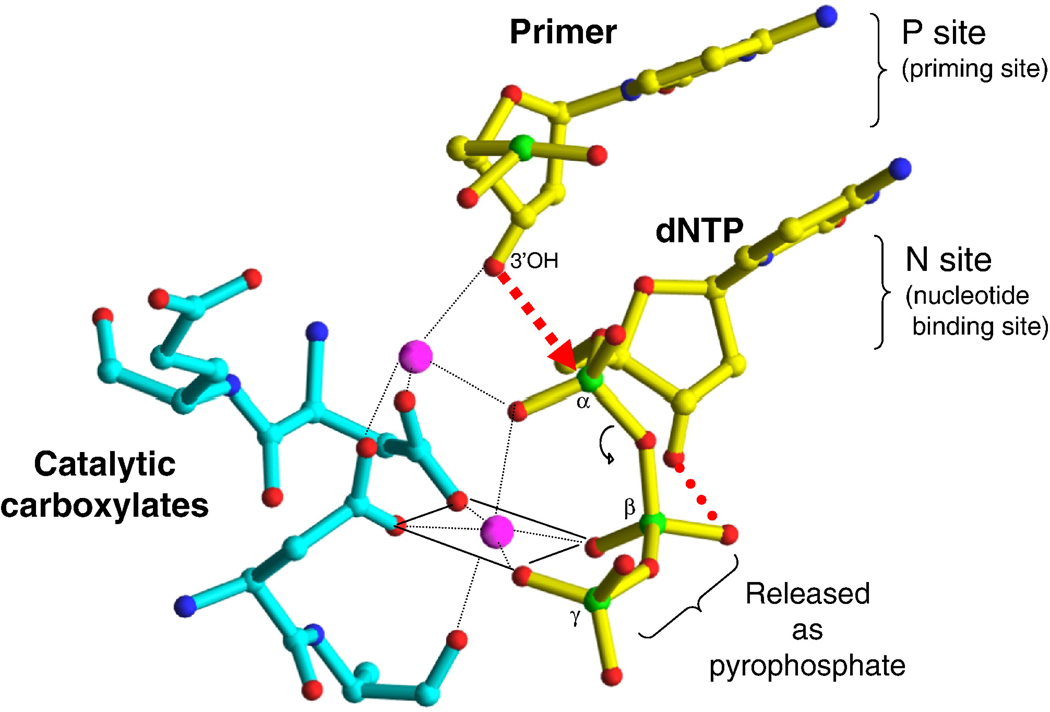

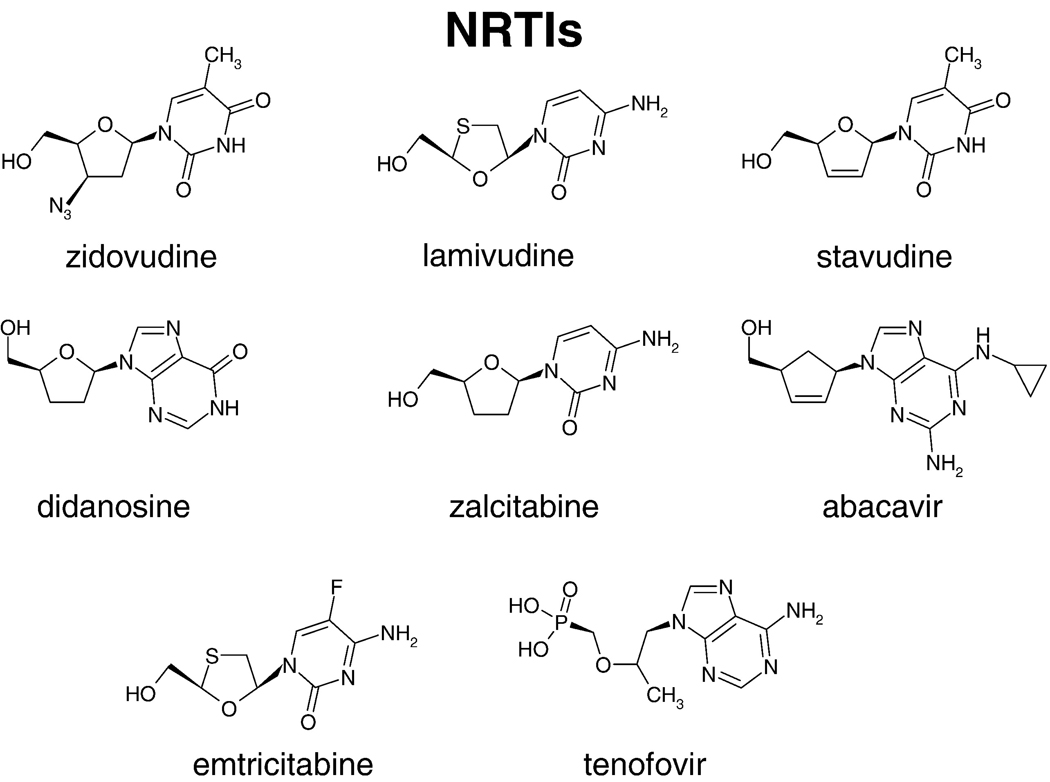
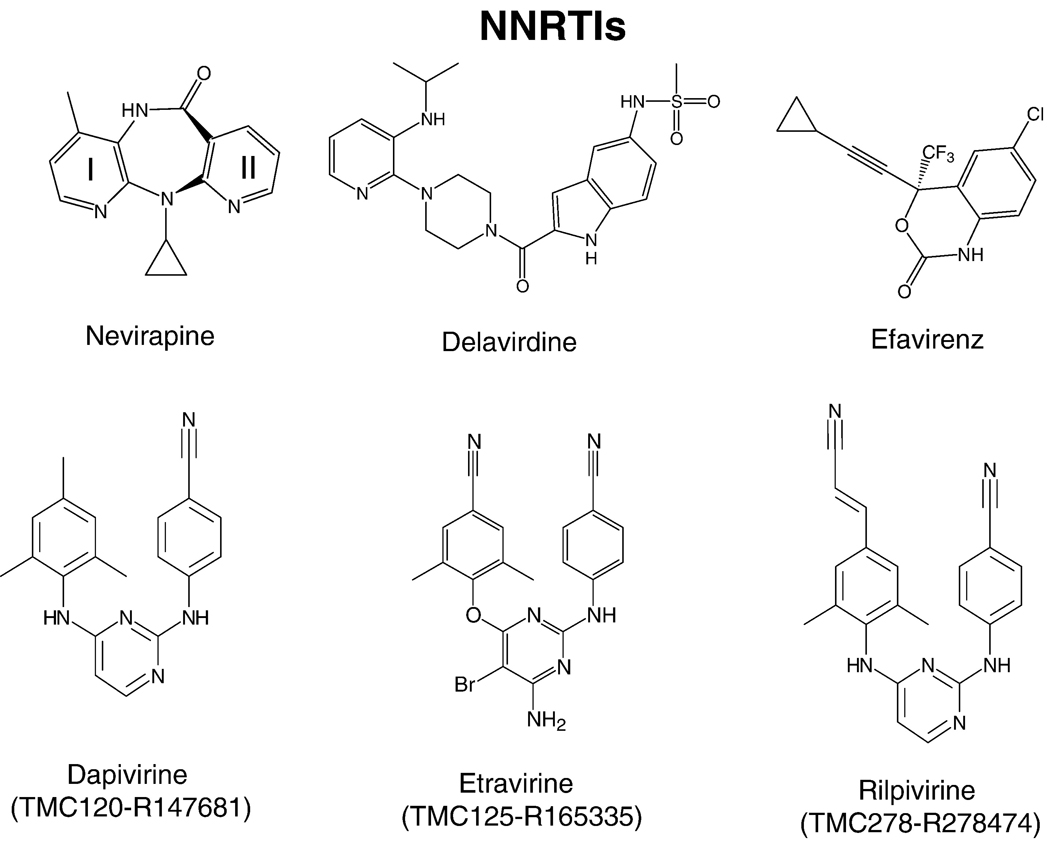
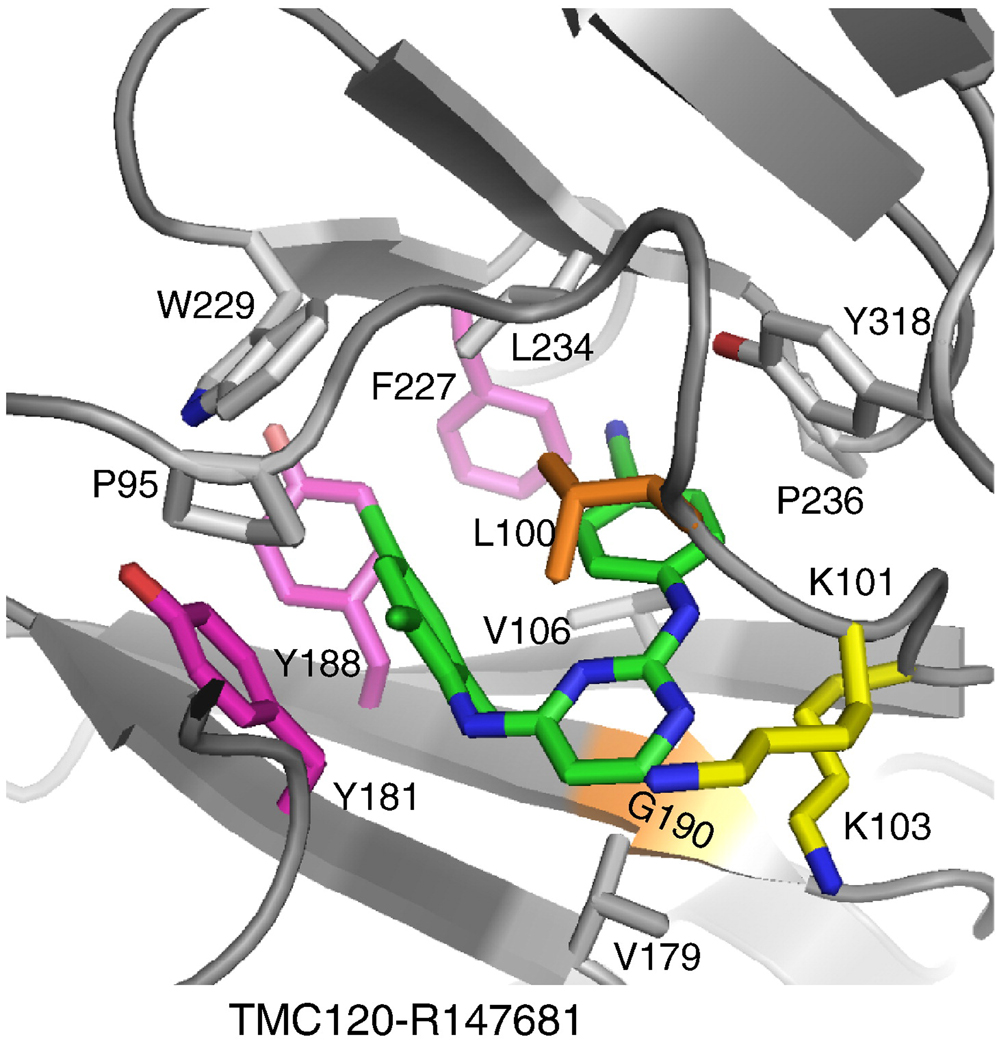
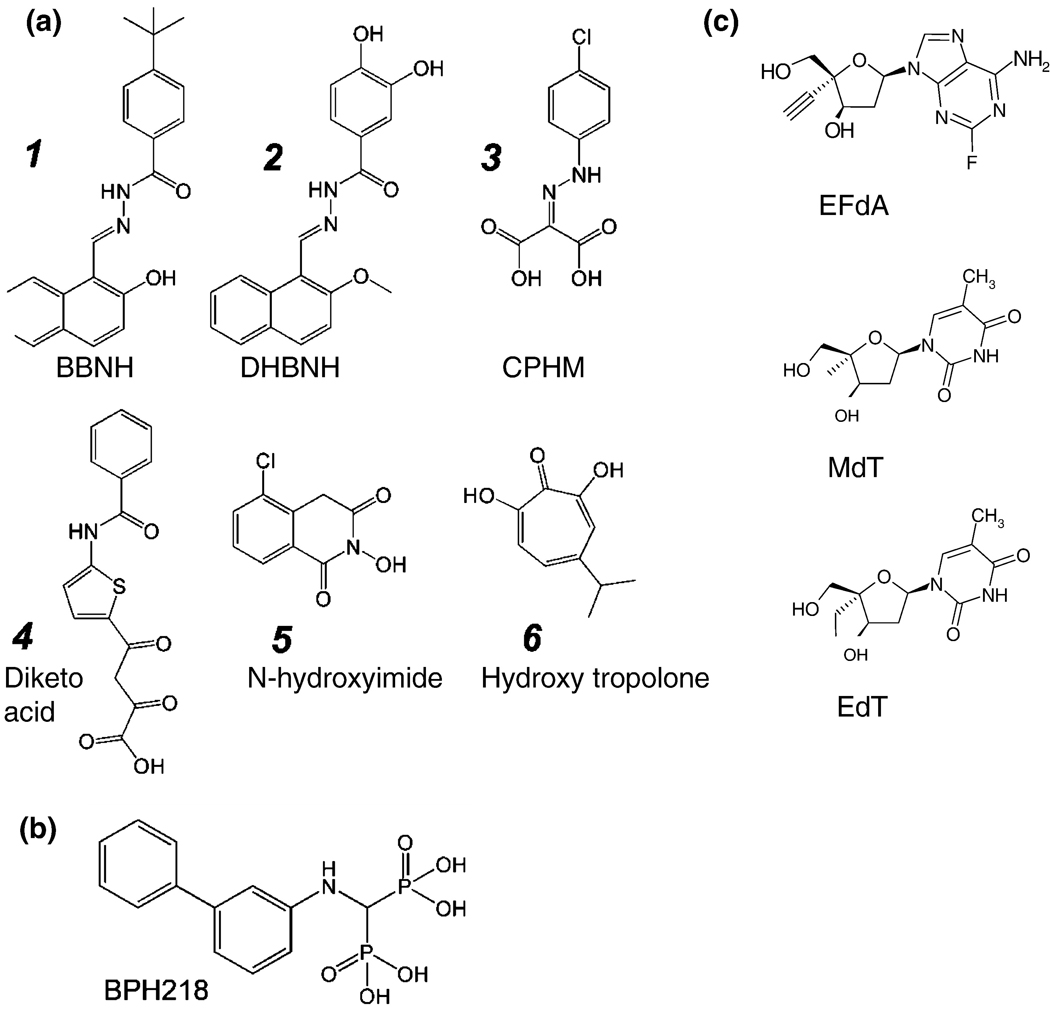
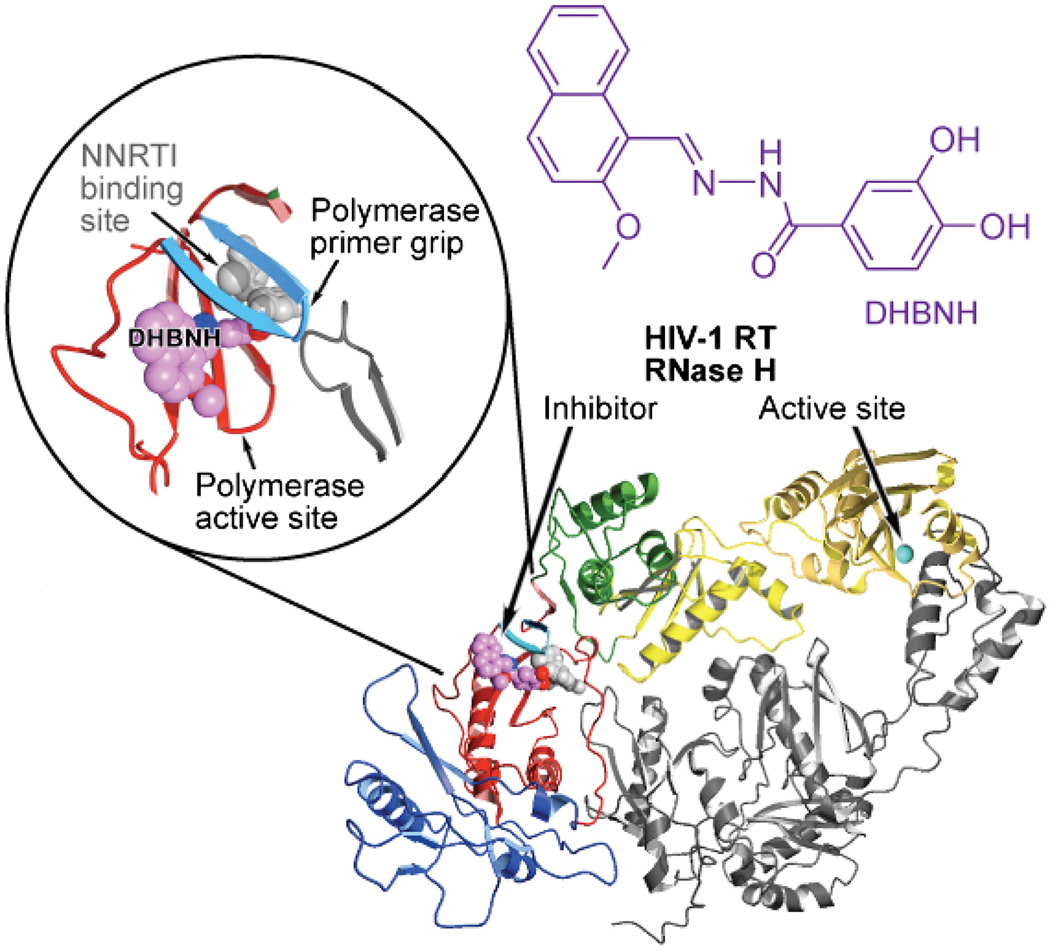
References
-
- Coffin JM, Hughes SH, Varmus HE. Retroviruses. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1997. - PubMed
-
- Coffin JM. HIV population dynamics in vivo: implications for genetic variation, pathogenesis, and therapy. Science. 1995;267:483–489. - PubMed
-
- di Marzo Veronese F, Copeland TD, DeVico AL, Rahman R, Oroszlan S, Gallo RC, Sarngadharan MG. Characterization of highly immunogenic p66/p51 as the reverse transcriptase of HTLV-III/LAV. Science. 1986;231:1289–1291. - PubMed
-
- Lowe DM, Aitken A, Bradley C, Darby GK, Larder BA, Powell KL, Purifoy DJ, Tisdale M, Stammers DK. HIV-1 reverse transcriptase: crystallization and analysis of domain structure by limited proteolysis. Biochemistry. 1988;27:8884–8889. - PubMed
-
- Kohlstaedt LA, Wang J, Friedman JM, Rice PA, Steitz TA. Crystal structure at 3.5 A resolution of HIV-1 reverse transcriptase complexed with an inhibitor. Science. 1992;256:1783–1790. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AI074389/AI/NIAID NIH HHS/United States
- U19 AI073975/AI/NIAID NIH HHS/United States
- R01 AI027690/AI/NIAID NIH HHS/United States
- R01 AI060452/AI/NIAID NIH HHS/United States
- P01 GM066671/GM/NIGMS NIH HHS/United States
- AI73975/AI/NIAID NIH HHS/United States
- AI074389/AI/NIAID NIH HHS/United States
- AI060452/AI/NIAID NIH HHS/United States
- AI076119/AI/NIAID NIH HHS/United States
- R37 AI076119/AI/NIAID NIH HHS/United States
- R01 AI076119/AI/NIAID NIH HHS/United States
- P01 GM 066671/GM/NIGMS NIH HHS/United States
- R37 AI027690/AI/NIAID NIH HHS/United States
- R33 AI079801/AI/NIAID NIH HHS/United States
- AI079801/AI/NIAID NIH HHS/United States
- R21 AI079801/AI/NIAID NIH HHS/United States
- ImNIH/Intramural NIH HHS/United States
- AI27690/AI/NIAID NIH HHS/United States
- AI07980/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
