

Proteomic and metabolomic analysis of cardioprotection: Interplay between protein kinase C epsilon and delta in regulating glucose metabolism of murine hearts
- PMID: 19027023
- PMCID: PMC3661410
- DOI: 10.1016/j.yjmcc.2008.10.008
Proteomic and metabolomic analysis of cardioprotection: Interplay between protein kinase C epsilon and delta in regulating glucose metabolism of murine hearts
Abstract
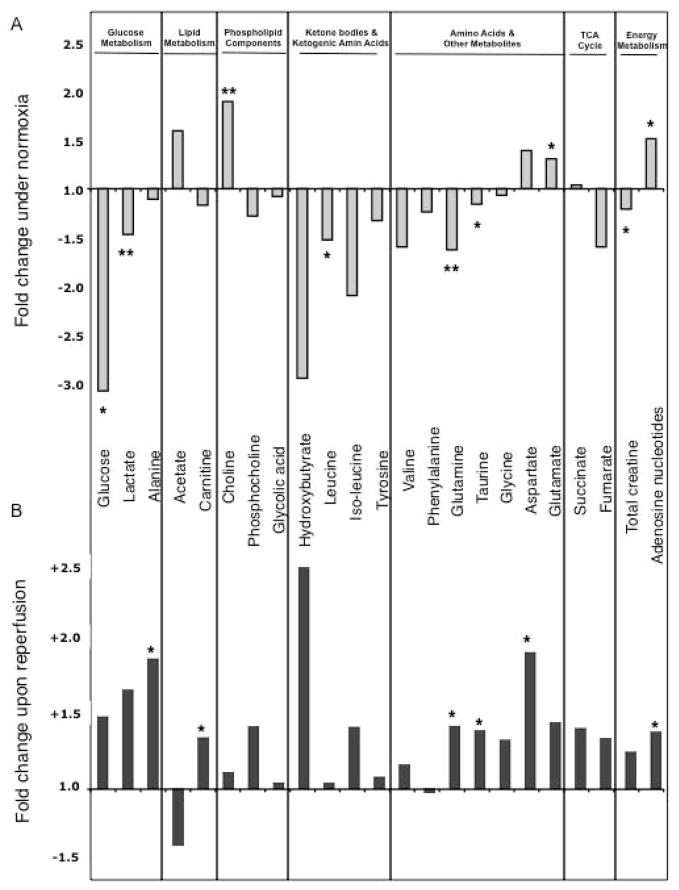
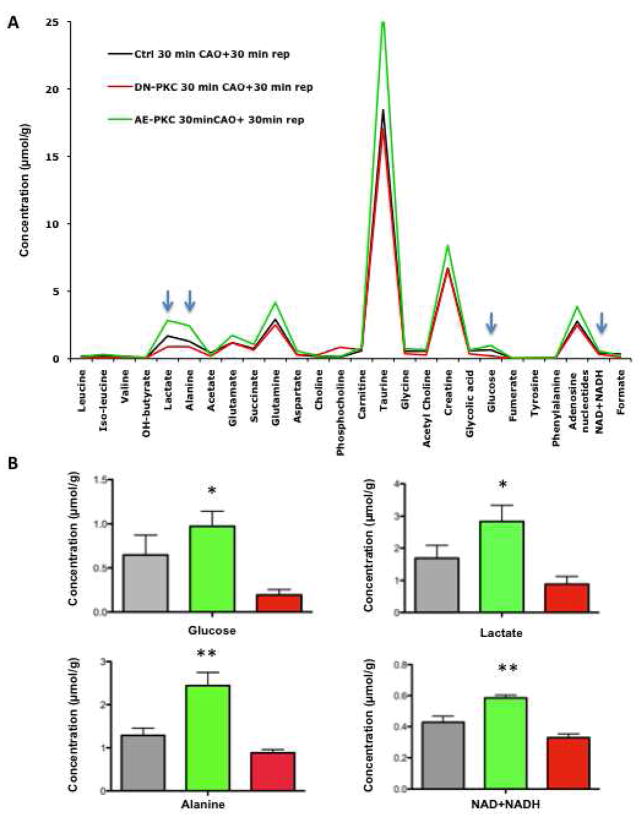
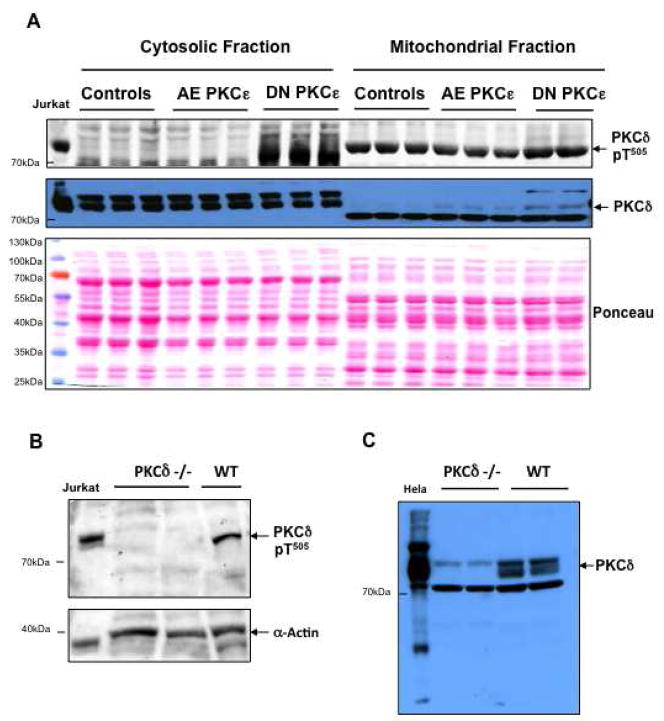
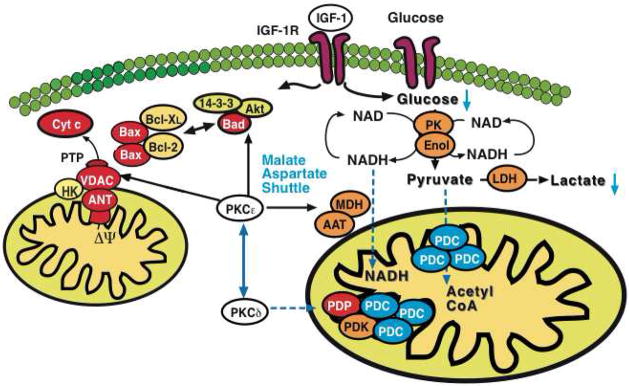
We applied a combined proteomic and metabolomic approach to obtain novel mechanistic insights in PKCvarepsilon-mediated cardioprotection. Mitochondrial and cytosolic proteins from control and transgenic hearts with constitutively active or dominant negative PKCvarepsilon were analyzed using difference in-gel electrophoresis (DIGE). Among the differentially expressed proteins were creatine kinase, pyruvate kinase, lactate dehydrogenase, and the cytosolic isoforms of aspartate amino transferase and malate dehydrogenase, the two enzymatic components of the malate aspartate shuttle, which are required for the import of reducing equivalents from glycolysis across the inner mitochondrial membrane. These enzymatic changes appeared to be dependent on PKCvarepsilon activity, as they were not observed in mice expressing inactive PKCvarepsilon. High-resolution proton nuclear magnetic resonance ((1)H-NMR) spectroscopy confirmed a pronounced effect of PKCvarepsilon activity on cardiac glucose and energy metabolism: normoxic hearts with constitutively active PKCvarepsilon had significantly lower concentrations of glucose, lactate, glutamine and creatine, but higher levels of choline, glutamate and total adenosine nucleotides. Moreover, the depletion of cardiac energy metabolites was slower during ischemia/reperfusion injury and glucose metabolism recovered faster upon reperfusion in transgenic hearts with active PKCvarepsilon. Notably, inhibition of PKCvarepsilon resulted in compensatory phosphorylation and mitochondrial translocation of PKCdelta. Taken together, our findings are the first evidence that PKCvarepsilon activity modulates cardiac glucose metabolism and provide a possible explanation for the synergistic effect of PKCdelta and PKCvarepsilon in cardioprotection.
Figures
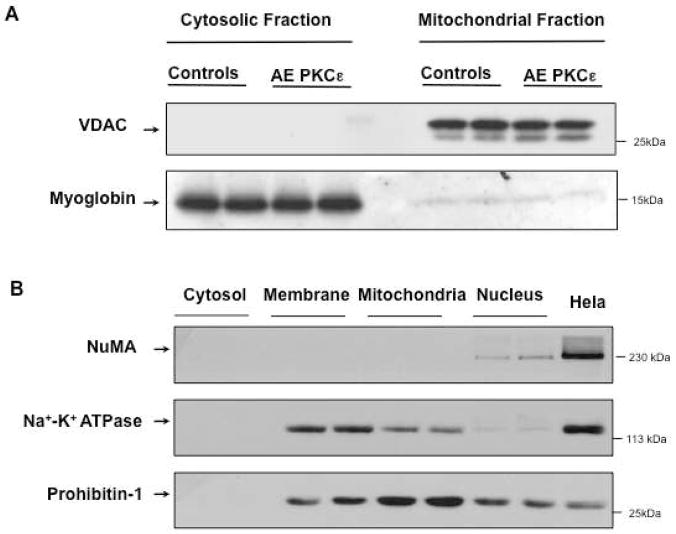
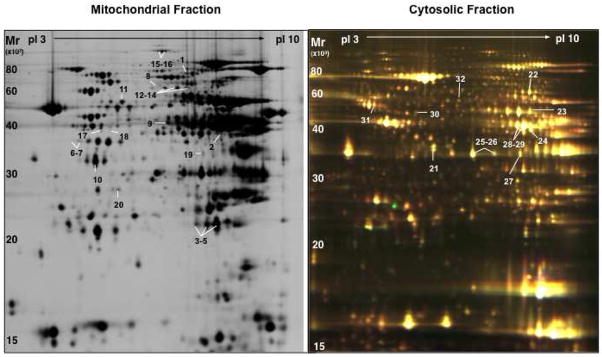
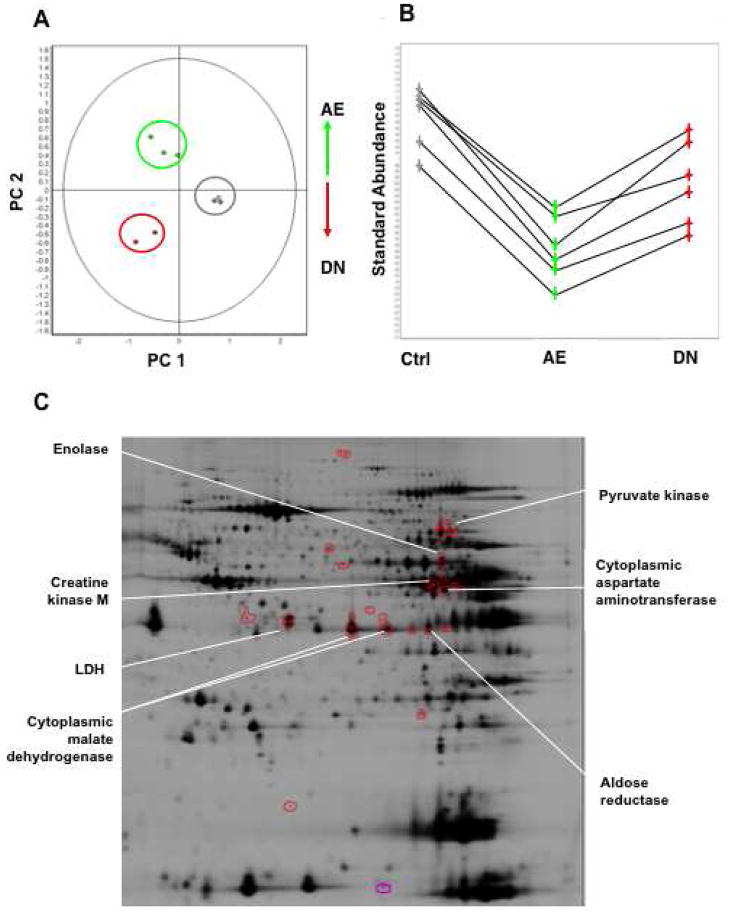
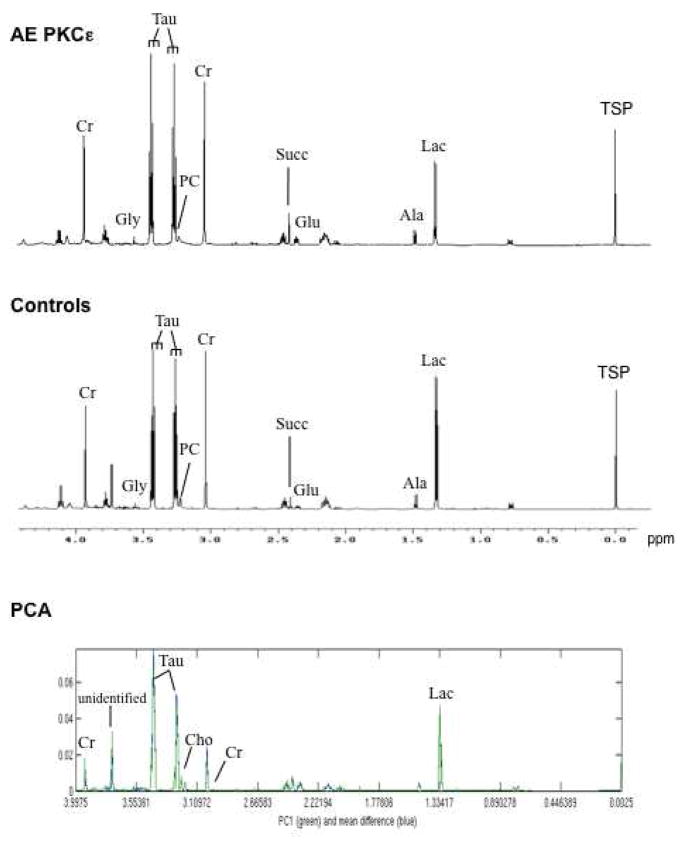
References
-
- Mackay K, Mochly-Rosen D. Localization, anchoring, and functions of protein kinase C isozymes in the heart. J Mol Cell Cardiol. 2001 Jul;33(7):1301–7. - PubMed
-
- Hahn HS, Yussman MG, Toyokawa T, Marreez Y, Barrett TJ, Hilty KC, et al. Ischemic protection and myofibrillar cardiomyopathy: dose-dependent effects of in vivo deltaPKC inhibition. Circ Res. 2002 Oct 18;91(8):741–8. - PubMed
-
- Inagaki K, Hahn HS, Dorn GW, 2nd, Mochly-Rosen D. Additive Protection of the Ischemic Heart Ex Vivo by Combined Treatment With {delta}-Protein Kinase C Inhibitor and {epsilon}-Protein Kinase C Activator. Circulation. 2003 Jul 14; - PubMed
-
- Murriel CL, Churchill E, Inagaki K, Szweda LI, Mochly-Rosen D. Protein Kinase C{delta} Activation Induces Apoptosis in Response to Cardiac Ischemia and Reperfusion Damage: A MECHANISM INVOLVING BAD AND THE MITOCHONDRIA. J Biol Chem. 2004 Nov 12;279(46):47985–91. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- HL-80111/HL/NHLBI NIH HHS/United States
- P01 HL080111/HL/NHLBI NIH HHS/United States
- HL63901/HL/NHLBI NIH HHS/United States
- R01 HL063901/HL/NHLBI NIH HHS/United States
- R01 HL065431/HL/NHLBI NIH HHS/United States
- FS/08/002/24537/BHF_/British Heart Foundation/United Kingdom
- HL-76526/HL/NHLBI NIH HHS/United States
- HL65431/HL/NHLBI NIH HHS/United States
- R01 HL080691/HL/NHLBI NIH HHS/United States
- HL 80691/HL/NHLBI NIH HHS/United States
- R21 HL076526/HL/NHLBI NIH HHS/United States
- R37 HL063901/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
