

The capsid proteins of a large, icosahedral dsDNA virus
- PMID: 19027752
- PMCID: PMC2911444
- DOI: 10.1016/j.jmb.2008.11.002
The capsid proteins of a large, icosahedral dsDNA virus
Abstract
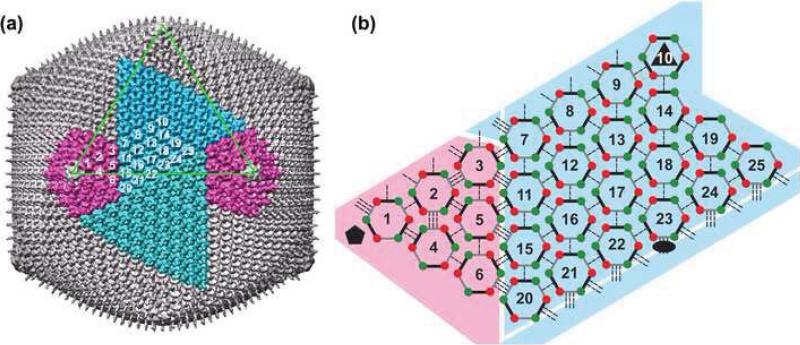
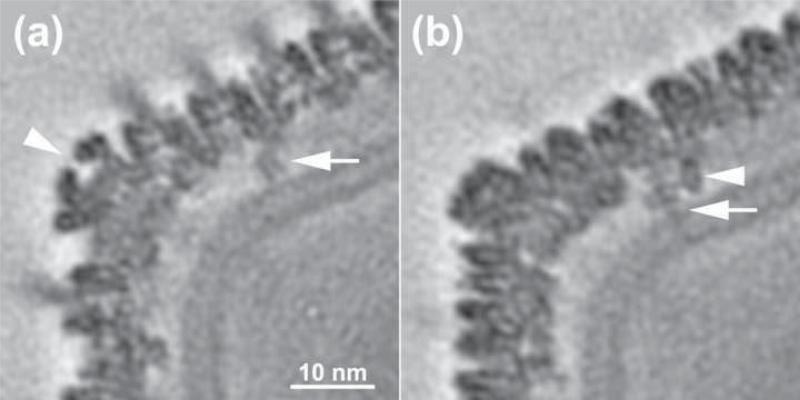
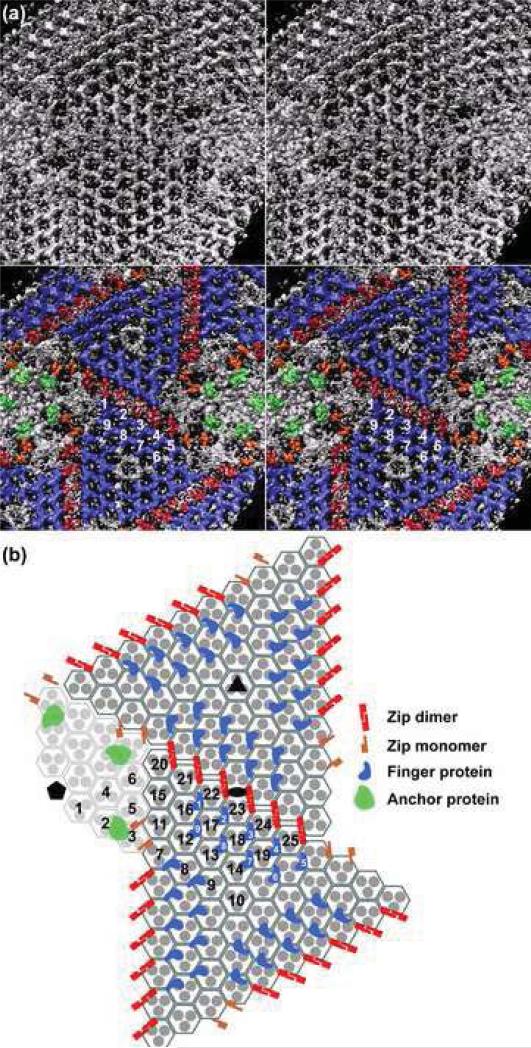
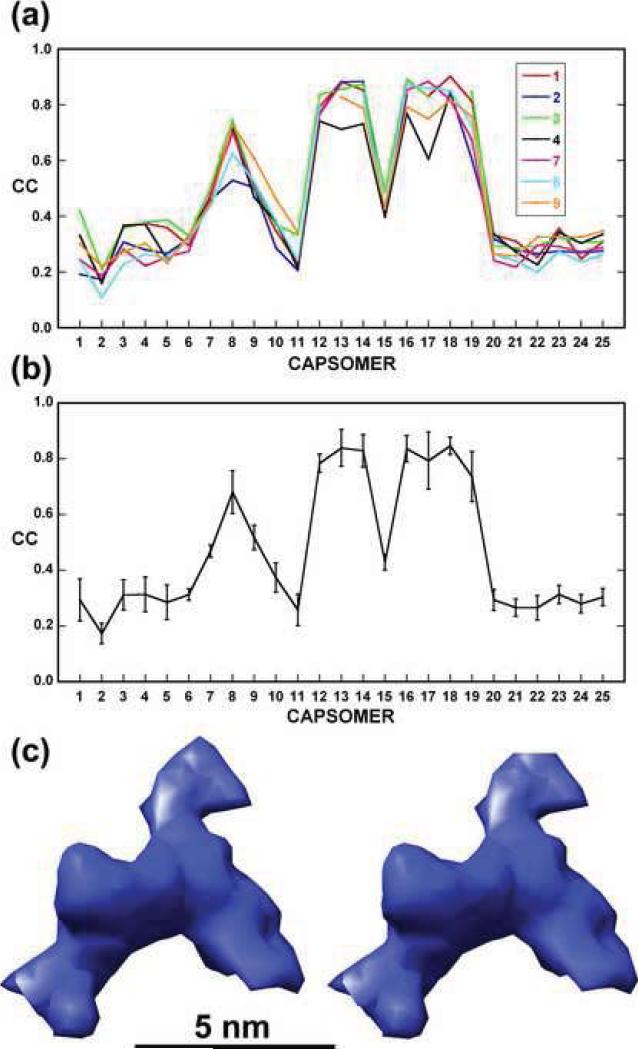
Chilo iridescent virus (CIV) is a large (approximately 1850 A diameter) insect virus with an icosahedral, T=147 capsid, a double-stranded DNA (dsDNA) genome, and an internal lipid membrane. The structure of CIV was determined to 13 A resolution by means of cryoelectron microscopy (cryoEM) and three-dimensional image reconstruction. A homology model of P50, the CIV major capsid protein (MCP), was built based on its amino acid sequence and the structure of the homologous Paramecium bursaria chlorella virus 1 Vp54 MCP. This model was fitted into the cryoEM density for each of the 25 trimeric CIV capsomers per icosahedral asymmetric unit. A difference map, in which the fitted CIV MCP capsomers were subtracted from the CIV cryoEM reconstruction, showed that there are at least three different types of minor capsid proteins associated with the capsomers outside the lipid membrane. "Finger" proteins are situated at many, but not all, of the spaces between three adjacent capsomers within each trisymmetron, and "zip" proteins are situated between sets of three adjacent capsomers at the boundary between neighboring trisymmetrons and pentasymmetrons. Based on the results of segmentation and density correlations, there are at least eight finger proteins and three dimeric and two monomeric zip proteins in one asymmetric unit of the CIV capsid. These minor proteins appear to stabilize the virus by acting as intercapsomer cross-links. One transmembrane "anchor" protein per icosahedral asymmetric unit, which extends from beneath one of the capsomers in the pentasymmetron to the internal leaflet of the lipid membrane, may provide additional stabilization for the capsid. These results are consistent with the observations for other large, icosahedral dsDNA viruses that also utilize minor capsid proteins for stabilization and for determining their assembly.
Figures

References
-
- Van Etten JL, Graves MV, MÜller DG, Boland W, Delaroque N. Phycodnaviridae - large DNA algal viruses. Arch. Virology. 2002;147:1479–1516. - PubMed
-
- La Scola B, Audic S, Robert C, Jungang L, de Lamballerie X, Drancourt M, Birtles R, Claverie JM, Raoult D. A giant virus in amoebae. Science. 2003;299:2033. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
