

The basolateral amygdala modulates specific sensory memory representations in the cerebral cortex
- PMID: 19028592
- PMCID: PMC3635825
- DOI: 10.1016/j.nlm.2008.10.010
The basolateral amygdala modulates specific sensory memory representations in the cerebral cortex
Abstract
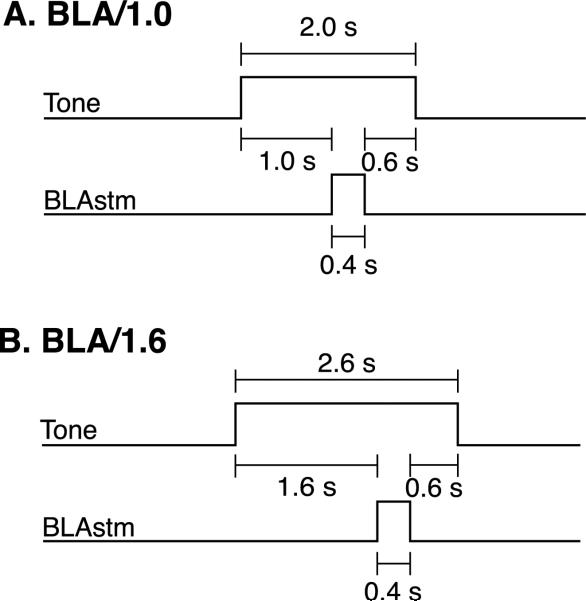
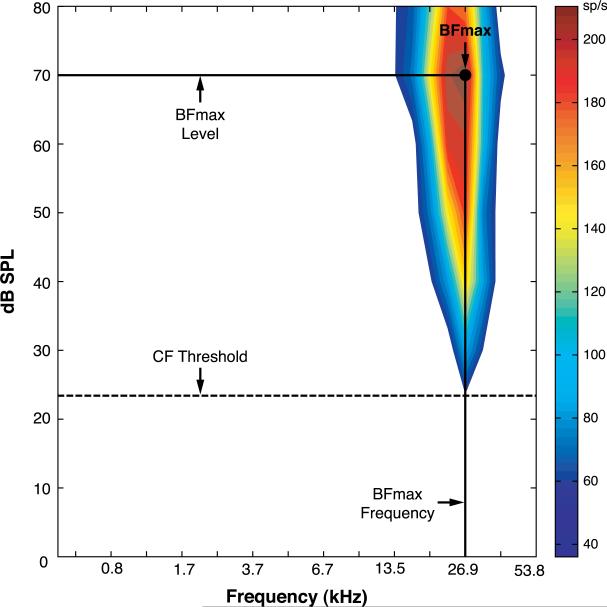
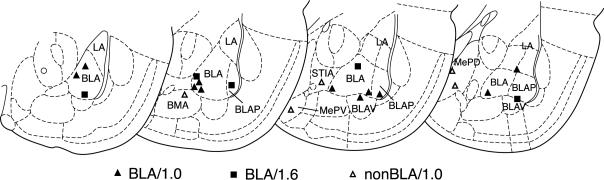
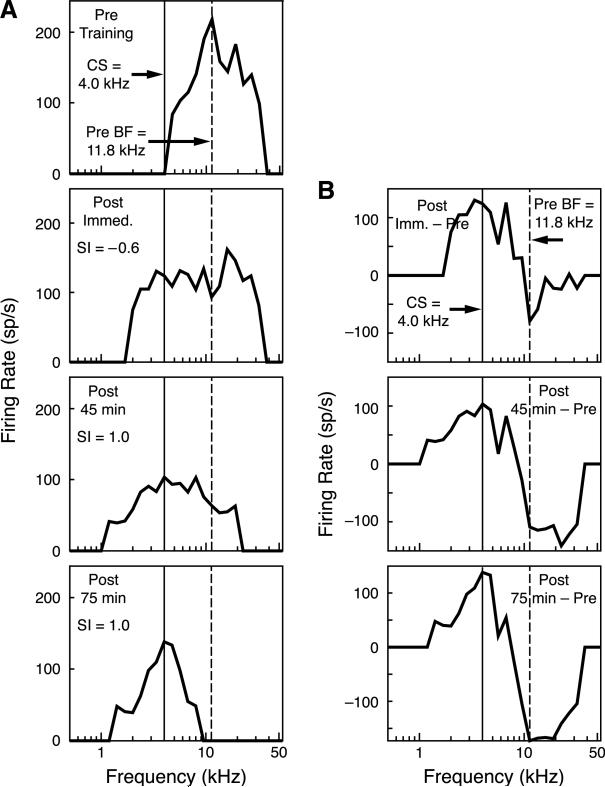
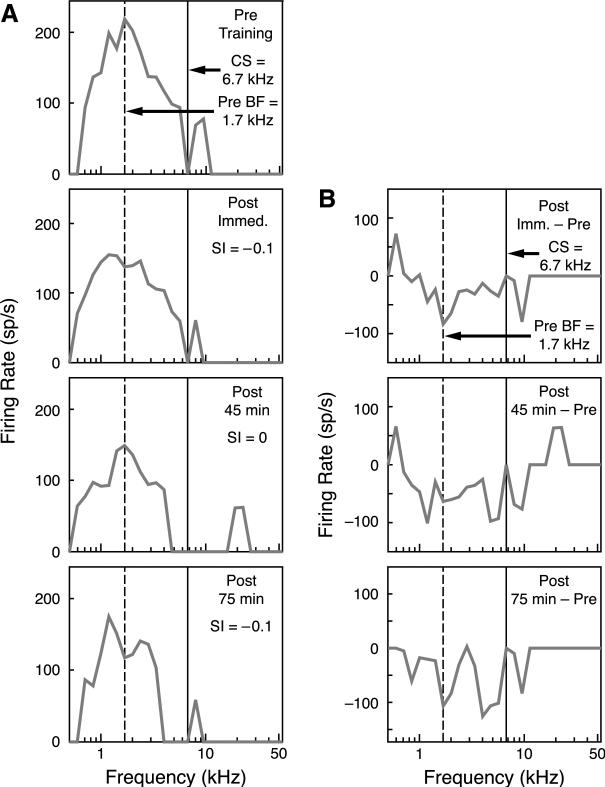
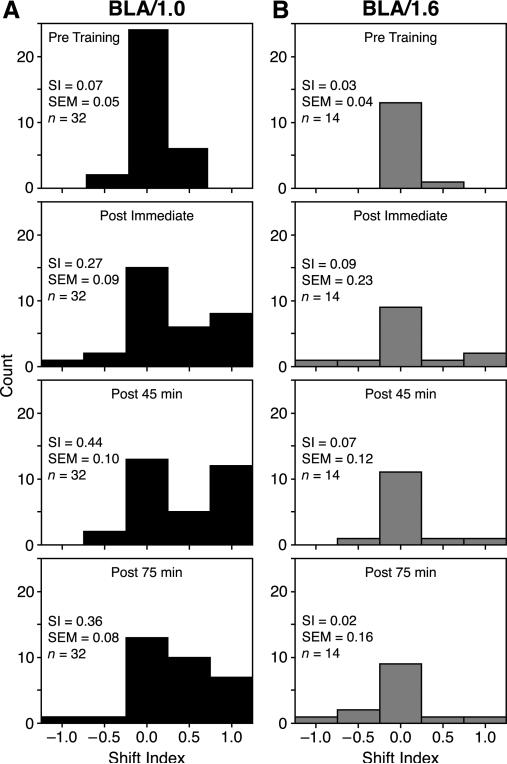
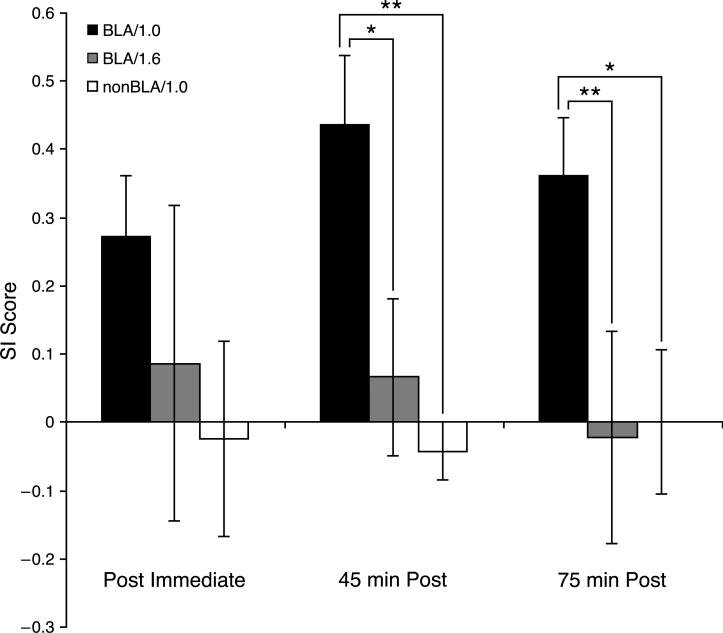
Stress hormones released by an experience can modulate memory strength via the basolateral amygdala, which in turn acts on sites of memory storage such as the cerebral cortex [McGaugh, J. L. (2004). The amygdala modulates the consolidation of memories of emotionally arousing experiences. Annual Review of Neuroscience, 27, 1-28]. Stimuli that acquire behavioral importance gain increased representation in the cortex. For example, learning shifts the tuning of neurons in the primary auditory cortex (A1) to the frequency of a conditioned stimulus (CS), and the greater the level of CS importance, the larger the area of representational gain [Weinberger, N. M. (2007). Associative representational plasticity in the auditory cortex: A synthesis of two disciplines. Learning & Memory, 14(1-2), 1-16]. The two lines of research suggest that BLA strengthening of memory might be accomplished in part by increasing the representation of an environmental stimulus. The present study investigated whether stimulation of the BLA can affect cortical memory representations. In male Sprague-Dawley rats studied under urethane general anesthesia, frequency receptive fields were obtained from A1 before and up to 75min after the pairing of a tone with BLA stimulation (BLAstm: 100 trials, 400ms, 100Hz, 400microA [+/-16.54]). Tone started before and continued after BLAstm. Group BLA/1.0 (n=16) had a 1s CS-BLAstm interval while Group BLA/1.6 (n=5) has a 1.6s interval. The BLA/1.0 group did develop specific tuning shifts toward and to the CS, which could change frequency tuning by as much as two octaves. Moreover, its shifts increased over time and were enduring, lasting 75min. However, group BLA/1.6 did not develop tuning shifts, indicating that precise CS-BLAstm timing is important in the anesthetized animal. Further, training in the BLA/1.0 paradigm but stimulating outside of the BLA did not produce tuning shifts. These findings demonstrate that the BLA is capable of exerting highly specific, enduring, learning-related modifications of stimulus representation in the cerebral cortex. These findings suggest that the ability of the BLA to alter specific cortical representations may underlie, at least in part, the modulatory influence of BLA activity on strengthening long-term memory.
Figures
References
-
- Bakin JS, Lepan B, Weinberger NM. Sensitization induced receptive field plasticity in the auditory cortex is independent of CS-modality. Brain Research. 1992;577(2):226–235. - PubMed
-
- Bakin JS, Weinberger NM. Classical conditioning induces CS-specific receptive field plasticity in the auditory cortex of the guinea pig. Brain Research. 1990;536(1–2):271–286. - PubMed
-
- Bao S, Chan VT, Merzenich MM. Cortical remodelling induced by activity of ventral tegmental dopamine neurons. Nature. 2001;412(6842):79–83. - PubMed
-
- Bergado JA, Rojas Y, Capdevila V, González O, Almaguer-Melian W. Stimulation of the basolateral amygdala improves the acquisition of a motor skill. Restorative Neurology and Neuroscience. 2006;24(2):115–121. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
