

Retinol Esterification by DGAT1 Is Essential for Retinoid Homeostasis in Murine Skin
- PMID: 19028692
- PMCID: PMC2640966
- DOI: 10.1074/jbc.M807503200
Retinol Esterification by DGAT1 Is Essential for Retinoid Homeostasis in Murine Skin
Abstract
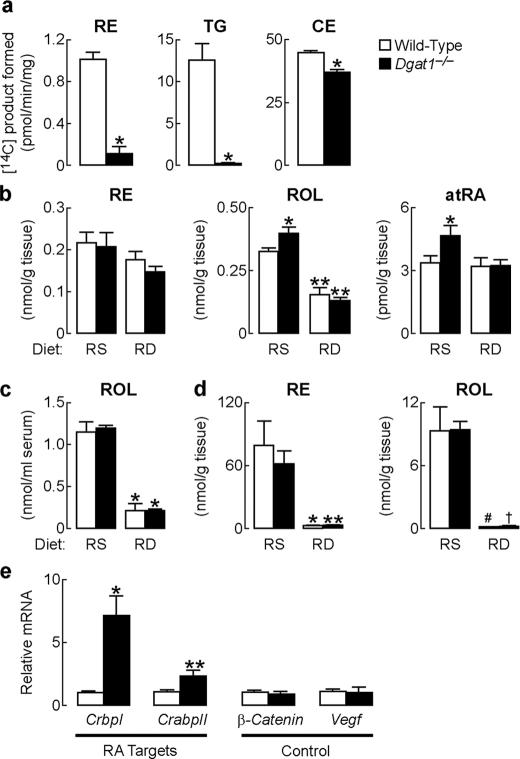
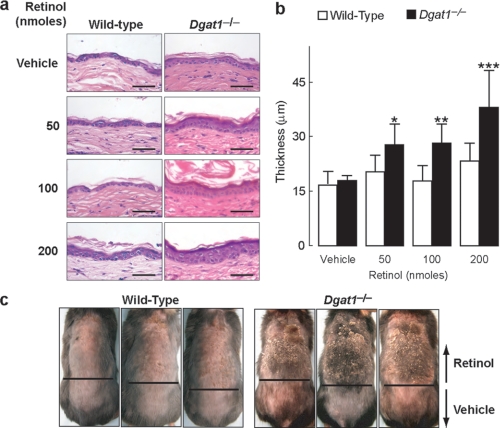
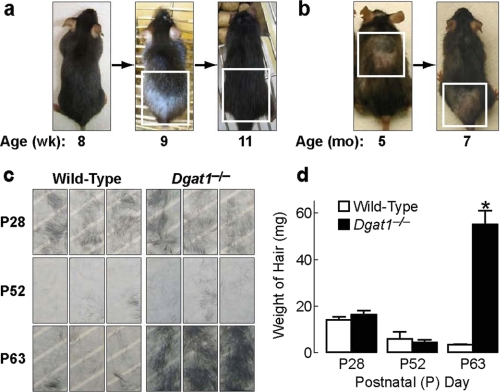
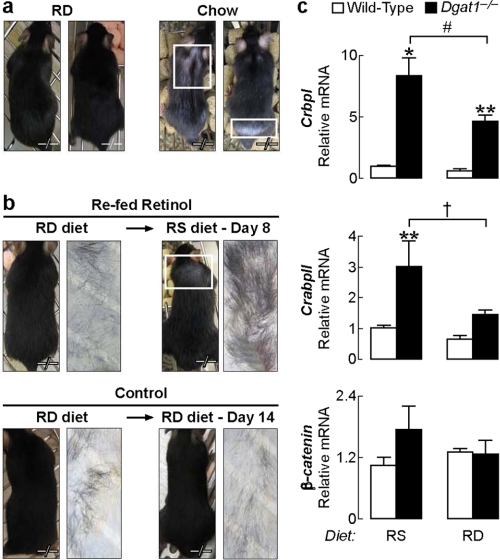
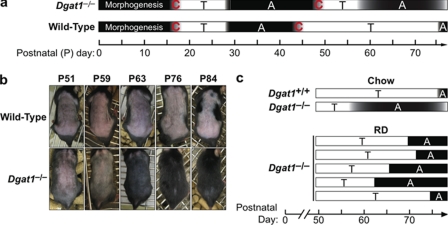
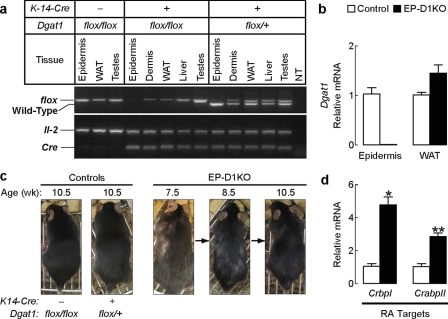
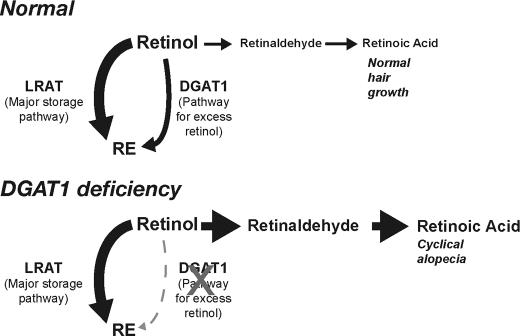
Retinoic acid (RA) is a potent signaling molecule that is essential for many biological processes, and its levels are tightly regulated by mechanisms that are only partially understood. The synthesis of RA from its precursor retinol (vitamin A) is an important regulatory mechanism. Therefore, the esterification of retinol with fatty acyl moieties to generate retinyl esters, the main storage form of retinol, may also regulate RA levels. Here we show that the neutral lipid synthesis enzyme acyl-CoA:diacylglycerol acyltransferase 1 (DGAT1) functions as the major acyl-CoA:retinol acyltransferase (ARAT) in murine skin. When dietary retinol is abundant, DGAT1 deficiency results in elevated levels of RA in skin and cyclical hair loss; both are prevented by dietary retinol deprivation. Further, DGAT1-deficient skin exhibits enhanced sensitivity to topically administered retinol. Deletion of the enzyme specifically in the epidermis causes alopecia, indicating that the regulation of RA homeostasis by DGAT1 is autonomous in the epidermis. These findings show that DGAT1 functions as an ARAT in the skin, where it acts to maintain retinoid homeostasis and prevent retinoid toxicity. Our findings may have implications for human skin or hair disorders treated with agents that modulate RA signaling.
Figures
References
-
- Balmer, J. E., and Blomhoff, R. (2002) J. Lipid Res. 43 1773-1808 - PubMed
-
- Mark, M., Ghyselinck, N. B., and Chambon, P. (2006) Annu. Rev. Pharmacol. Toxicol. 46 451-480 - PubMed
-
- Azais-Braesco, V., and Pascal, G. (2000) Am. J. Clin. Nutr. 71 1325S-1333S - PubMed
-
- Hansen, L. A., Sigman, C. C., Andreola, F., Ross, S. A., Kelloff, G. J., and De Luca, L. M. (2000) Carcinogenesis 21 1271-1279 - PubMed
-
- Napoli, J. L. (1999) Biochim. Biophys. Acta 1440 139-162 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
