

Functional UDP-xylose transport across the endoplasmic reticulum/Golgi membrane in a Chinese hamster ovary cell mutant defective in UDP-xylose Synthase
- PMID: 19028698
- PMCID: PMC2629108
- DOI: 10.1074/jbc.M804394200
Functional UDP-xylose transport across the endoplasmic reticulum/Golgi membrane in a Chinese hamster ovary cell mutant defective in UDP-xylose Synthase
Abstract
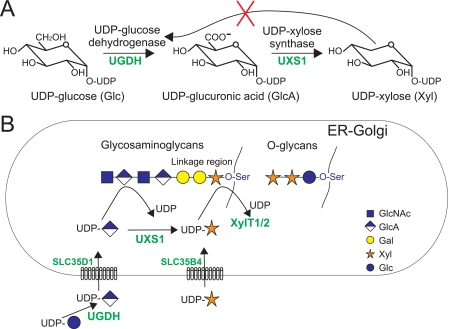
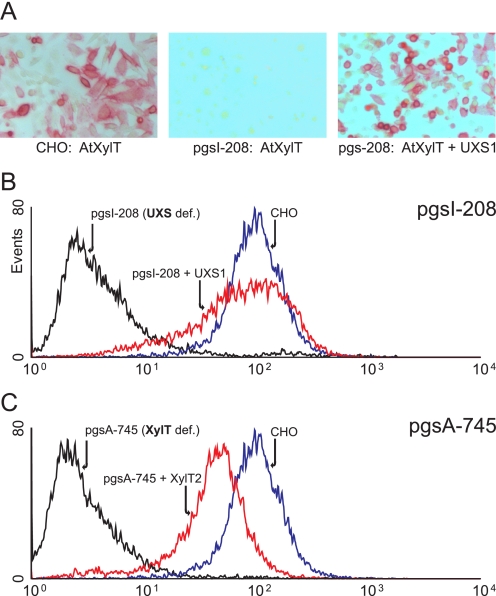
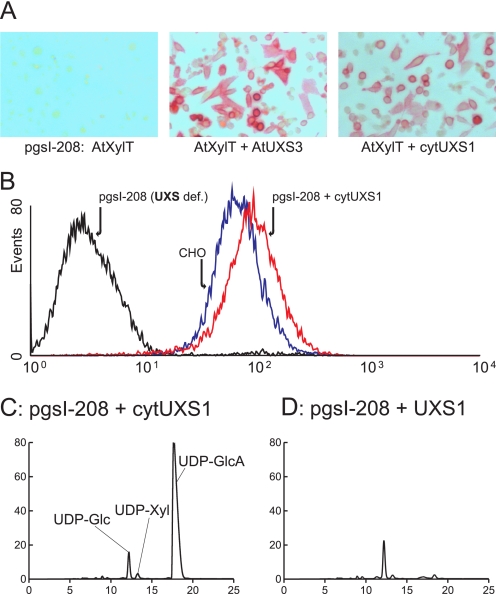
In mammals, xylose is found as the first sugar residue of the tetrasaccharide GlcAbeta1-3Galbeta1-3Galbeta1-4Xylbeta1-O-Ser, initiating the formation of the glycosaminoglycans heparin/heparan sulfate and chondroitin/dermatan sulfate. It is also found in the trisaccharide Xylalpha1-3Xylalpha1-3Glcbeta1-O-Ser on epidermal growth factor repeats of proteins, such as Notch. UDP-xylose synthase (UXS), which catalyzes the formation of the UDP-xylose substrate for the different xylosyltransferases through decarboxylation of UDP-glucuronic acid, resides in the endoplasmic reticulum and/or Golgi lumen. Since xylosylation takes place in these organelles, no obvious requirement exists for membrane transport of UDP-xylose. However, UDP-xylose transport across isolated Golgi membranes has been documented, and we recently succeeded with the cloning of a human UDP-xylose transporter (SLC25B4). Here we provide new evidence for a functional role of UDP-xylose transport by characterization of a new Chinese hamster ovary cell mutant, designated pgsI-208, that lacks UXS activity. The mutant fails to initiate glycosaminoglycan synthesis and is not capable of xylosylating Notch. Complementation was achieved by expression of a cytoplasmic variant of UXS, which proves the existence of a functional Golgi UDP-xylose transporter. A approximately 200 fold increase of UDP-glucuronic acid occurred in pgsI-208 cells, demonstrating a lack of UDP-xylose-mediated control of the cytoplasmically localized UDP-glucose dehydrogenase in the mutant. The data presented in this study suggest the bidirectional transport of UDP-xylose across endoplasmic reticulum/Golgi membranes and its role in controlling homeostasis of UDP-glucuronic acid and UDP-xylose production.
Figures


References
-
- Roden, L., and Armand, G. (1966) J. Biol. Chem. 241 65-70 - PubMed
-
- Hase, S., Kawabata, S., Nishimura, H., Takeya, H., Sueyoshi, T., Miyata, T., Iwanaga, S., Takao, T., Shimonishi, Y., and Ikenaka, T. (1988) J. Biochem. (Tokyo) 104 867-868 - PubMed
-
- Moloney, D. J., Shair, L. H., Lu, F. M., Xia, J., Locke, R., Matta, K. L., and Haltiwanger, R. S. (2000) J. Biol. Chem. 275 9604-9611 - PubMed
-
- Ponighaus, C., Ambrosius, M., Casanova, J. C., Prante, C., Kuhn, J., Esko, J. D., Kleesiek, K., and Götting, C. (2007) J. Biol. Chem. 282 5201-5206 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
