

Alterations in the endogenous ascorbic acid content affect flowering time in Arabidopsis
- PMID: 19028878
- PMCID: PMC2633856
- DOI: 10.1104/pp.108.132324
Alterations in the endogenous ascorbic acid content affect flowering time in Arabidopsis
Abstract
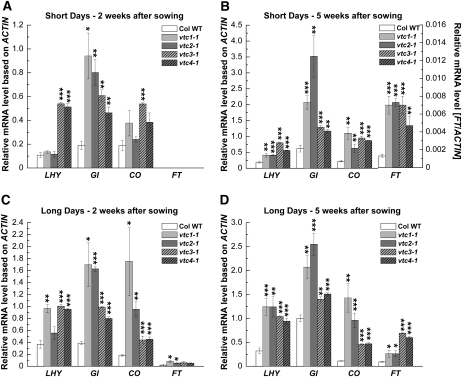
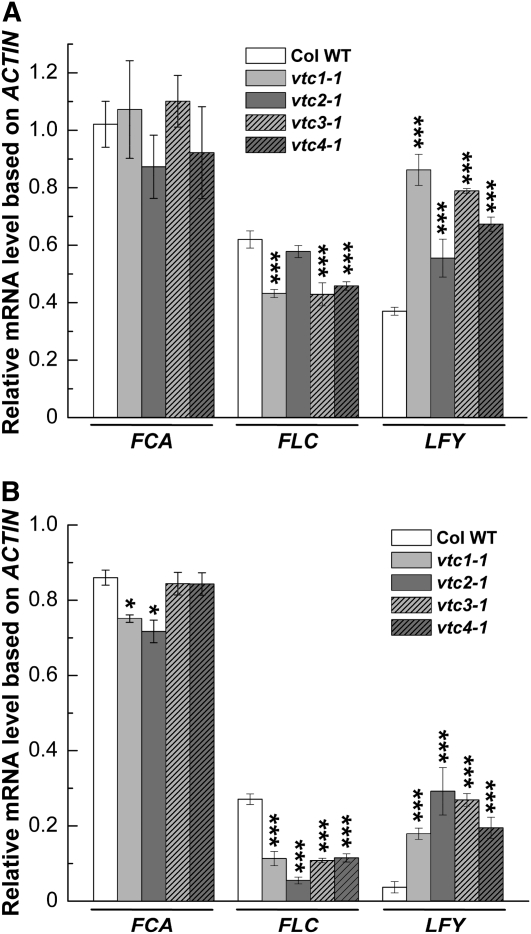
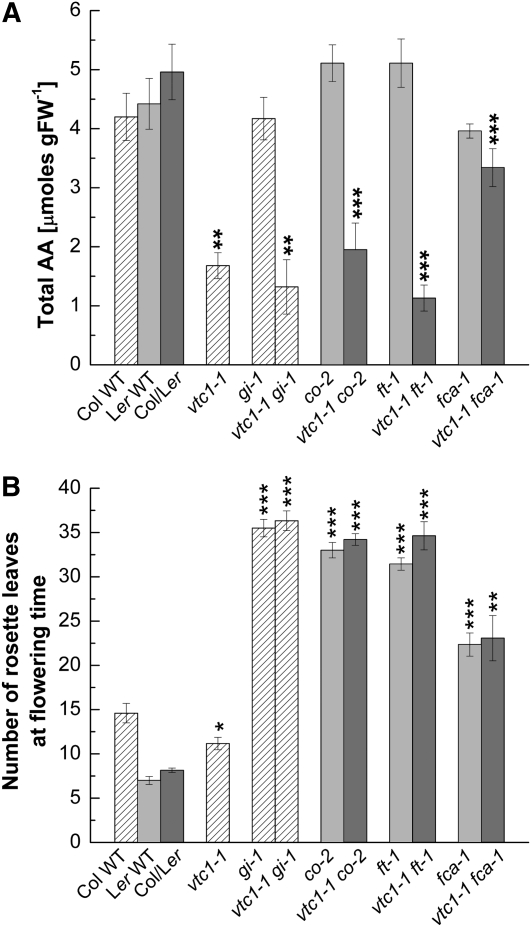
Ascorbic acid (AA) protects plants against abiotic stress. Previous studies suggested that this antioxidant is also involved in the control of flowering. To decipher how AA influences flowering time, we studied the four AA-deficient Arabidopsis (Arabidopsis thaliana) mutants vtc1-1, vtc2-1, vtc3-1, and vtc4-1 when grown under short and long days. These mutants flowered and senesced before the wild type irrespective of the photoperiod, a response that cannot simply be attributed to slightly elevated oxidative stress in the mutants. Transcript profiling of various flowering pathway genes revealed a correlation of altered mRNA levels and flowering time. For example, circadian clock and photoperiodic pathway genes were significantly higher in the vtc mutants than in the wild type under both short and long days, a result that is consistent with the early-flowering phenotype of the mutants. In contrast, when the AA content was artificially increased, flowering was delayed, which correlated with lower mRNA levels of circadian clock and photoperiodic pathway genes compared with plants treated with water. Similar observations were made for the autonomous pathway. Genetic analyses demonstrated that various photoperiodic and autonomous pathway mutants are epistatic to the vtc1-1 mutant. In conclusion, our transcript and genetic analyses suggest that AA acts upstream of the photoperiodic and autonomous pathways.
Figures


References
-
- Abe M, Kobayashi Y, Yamamoto S, Daimon Y, Yamaguchi A, Ikeda Y, Ichinoki H, Notaguchi M, Goto K, Araki T (2005) FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science 309 1052–1056 - PubMed
-
- Alabadi D, Oyama T, Yanovsky MJ, Harmon FG, Mas P, Kay SA (2001) Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science 293 880–883 - PubMed
-
- Araki T, Komeda Y (1993) Analysis of the role of the late-flowering locus, Gl, in the flowering of Arabidopsis thaliana. Plant J 3 231–239
-
- Arrigoni O, De Tullio MC (2000) The role of ascorbic acid in cell metabolism: between gene-directed functions and unpredictable chemical reactions. J Plant Physiol 157 481–488
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
