

Glycerol metabolism is important for cytotoxicity of Mycoplasma pneumoniae
- PMID: 19028882
- PMCID: PMC2632104
- DOI: 10.1128/JB.01103-08
Glycerol metabolism is important for cytotoxicity of Mycoplasma pneumoniae
Abstract
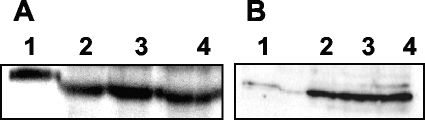
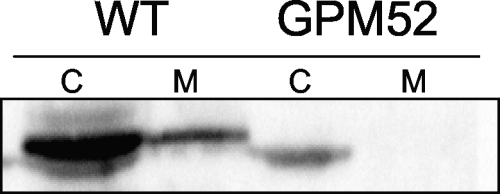
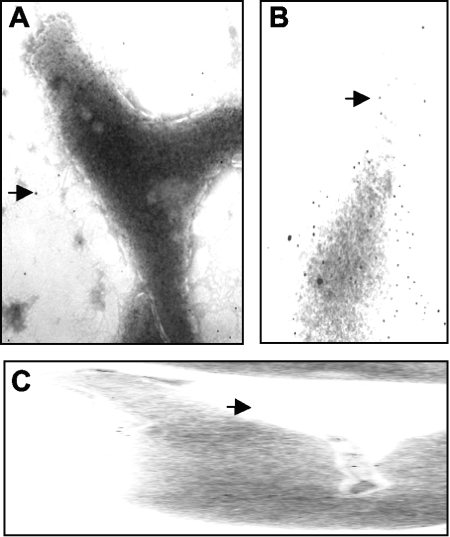
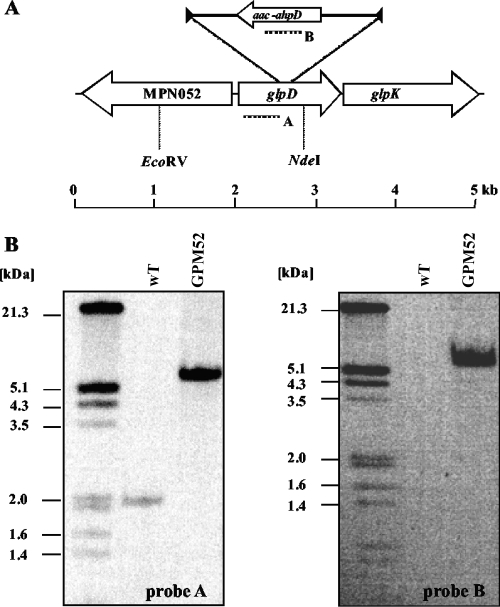
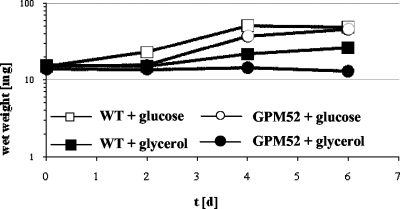
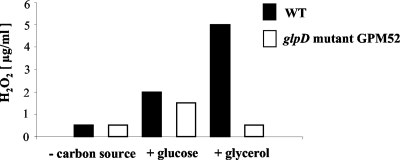
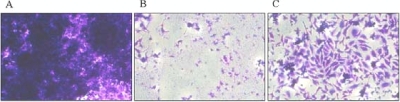
Glycerol is one of the few carbon sources that can be utilized by Mycoplasma pneumoniae. Glycerol metabolism involves uptake by facilitated diffusion, phosphorylation, and the oxidation of glycerol 3-phosphate to dihydroxyacetone phosphate, a glycolytic intermediate. We have analyzed the expression of the genes involved in glycerol metabolism and observed constitutive expression irrespective of the presence of glycerol or preferred carbon sources. Similarly, the enzymatic activity of glycerol kinase is not modulated by HPr-dependent phosphorylation. This lack of regulation is unique among the bacteria for which glycerol metabolism has been studied so far. Two types of enzymes catalyze the oxidation of glycerol 3-phosphate: oxidases and dehydrogenases. Here, we demonstrate that the enzyme encoded by the M. pneumoniae glpD gene is a glycerol 3-phosphate oxidase that forms hydrogen peroxide rather than NADH(2). The formation of hydrogen peroxide by GlpD is crucial for cytotoxic effects of M. pneumoniae. A glpD mutant exhibited a significantly reduced formation of hydrogen peroxide and a severely reduced cytotoxicity. Attempts to isolate mutants affected in the genes of glycerol metabolism revealed that only the glpD gene, encoding the glycerol 3-phosphate oxidase, is dispensable. In contrast, the glpF and glpK genes, encoding the glycerol facilitator and the glycerol kinase, respectively, are essential in M. pneumoniae. Thus, the enzymes of glycerol metabolism are crucial for the pathogenicity of M. pneumoniae but also for other essential, yet-to-be-identified functions in the M. pneumoniae cell.
Figures
References
-
- Carpousis, A. J. 2007. The RNA degradosome of Escherichia coli: an mRNA-degrading machine assembled on RNase E. Annu. Rev. Microbiol. 6171-87. - PubMed
-
- Darbon, E., P. Servant, S. Poncet, and J. Deutscher. 2002. Antitermination by GlpP, catabolite repression via CcpA and inducer exclusion triggered by P-GlpK dephosphorylation control Bacillus subtilis glpFK expression. Mol. Microbiol. 431039-1052. - PubMed
-
- Deutscher, J. 2008. The mechanisms of carbon catabolite repression in bacteria. Curr. Opin. Microbiol. 1187-93. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
