

Mutant telomeric repeats in yeast can disrupt the negative regulation of recombination-mediated telomere maintenance and create an alternative lengthening of telomeres-like phenotype
- PMID: 19029249
- PMCID: PMC2630680
- DOI: 10.1128/MCB.00423-08
Mutant telomeric repeats in yeast can disrupt the negative regulation of recombination-mediated telomere maintenance and create an alternative lengthening of telomeres-like phenotype
Abstract
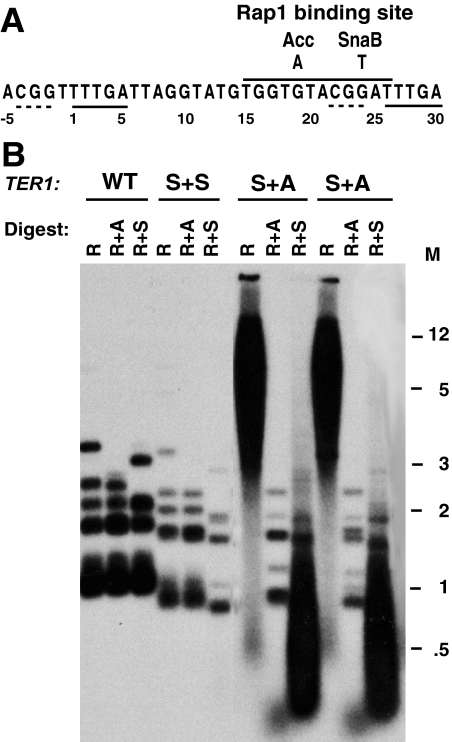
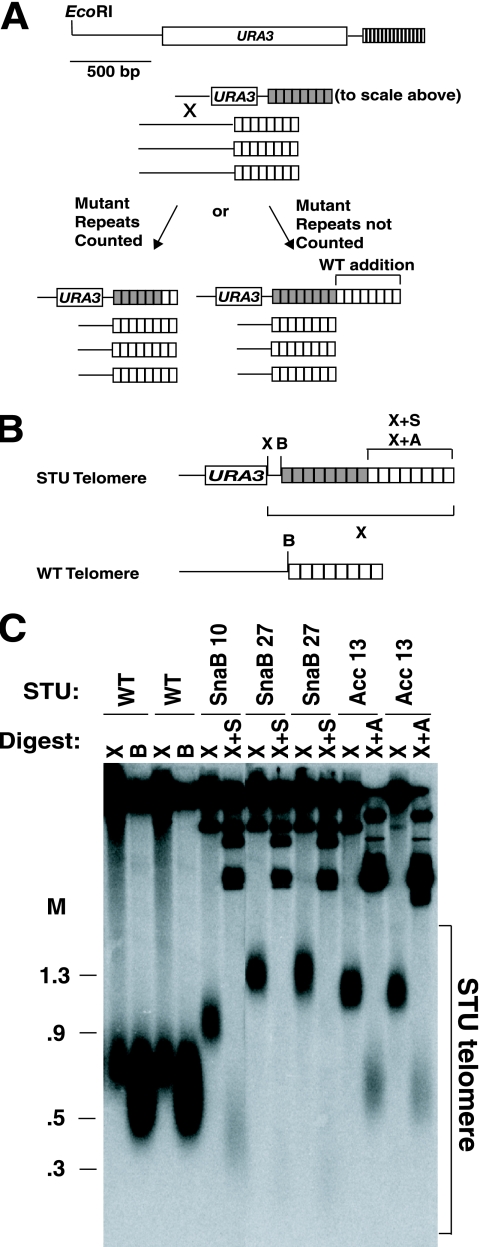
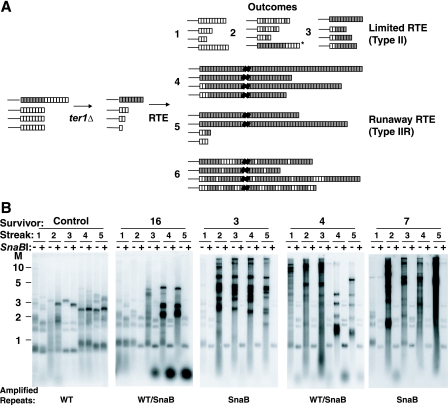
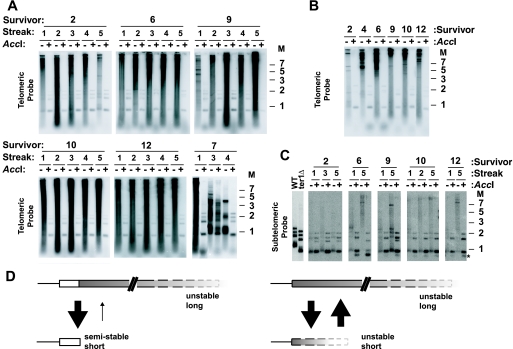
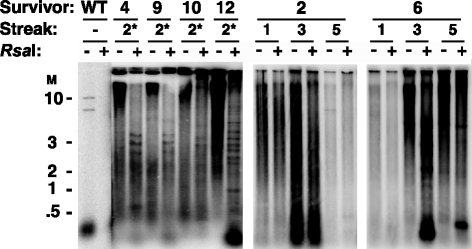
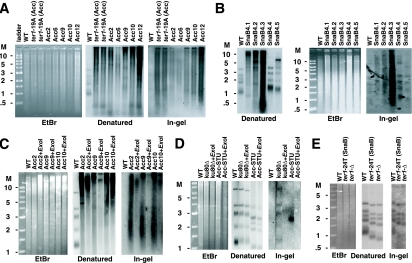
Some human cancers maintain telomeres using alternative lengthening of telomeres (ALT), a process thought to be due to recombination. In Kluyveromyces lactis mutants lacking telomerase, recombinational telomere elongation (RTE) is induced at short telomeres but is suppressed once telomeres are moderately elongated by RTE. Recent work has shown that certain telomere capping defects can trigger a different type of RTE that results in much more extensive telomere elongation that is reminiscent of human ALT cells. In this study, we generated telomeres composed of either of two types of mutant telomeric repeats, Acc and SnaB, that each alter the binding site for the telomeric protein Rap1. We show here that arrays of both types of mutant repeats present basally on a telomere were defective in negatively regulating telomere length in the presence of telomerase. Similarly, when each type of mutant repeat was spread to all chromosome ends in cells lacking telomerase, they led to the formation of telomeres produced by RTE that were much longer than those seen in cells with only wild-type telomeric repeats. The Acc repeats produced the more severe defect in both types of telomere maintenance, consistent with their more severe Rap1 binding defect. Curiously, although telomerase deletion mutants with telomeres composed of Acc repeats invariably showed extreme telomere elongation, they often also initially showed persistent very short telomeres with few or no Acc repeats. We suggest that these result from futile cycles of recombinational elongation and truncation of the Acc repeats from the telomeres. The presence of extensive 3' overhangs at mutant telomeres suggests that Rap1 may normally be involved in controlling 5' end degradation.
Figures
References
-
- Autexier, C., and N. F. Lue. 2006. The structure and function of telomerase reverse transcriptase. Annu. Rev. Biochem. 75493-517. - PubMed
-
- Baumann, P., and T. R. Cech. 2001. Pot1, the putative telomere end-binding protein in fission yeast and humans. Science 2921171-1175. - PubMed
-
- Beernink, H. T., K. Miller, A. Deshpande, P. Bucher, and J. P. Cooper. 2003. Telomere maintenance in fission yeast requires an Est1 ortholog. Curr. Biol. 13575-580. - PubMed
-
- Bertuch, A. A., and V. Lundblad. 2006. The maintenance and masking of chromosome termini. Curr. Opin. Cell Biol. 18247-253. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources