

The ribosome structure controls and directs mRNA entry, translocation and exit dynamics
- PMID: 19029596
- PMCID: PMC2907240
- DOI: 10.1088/1478-3975/5/4/046005
The ribosome structure controls and directs mRNA entry, translocation and exit dynamics
Abstract
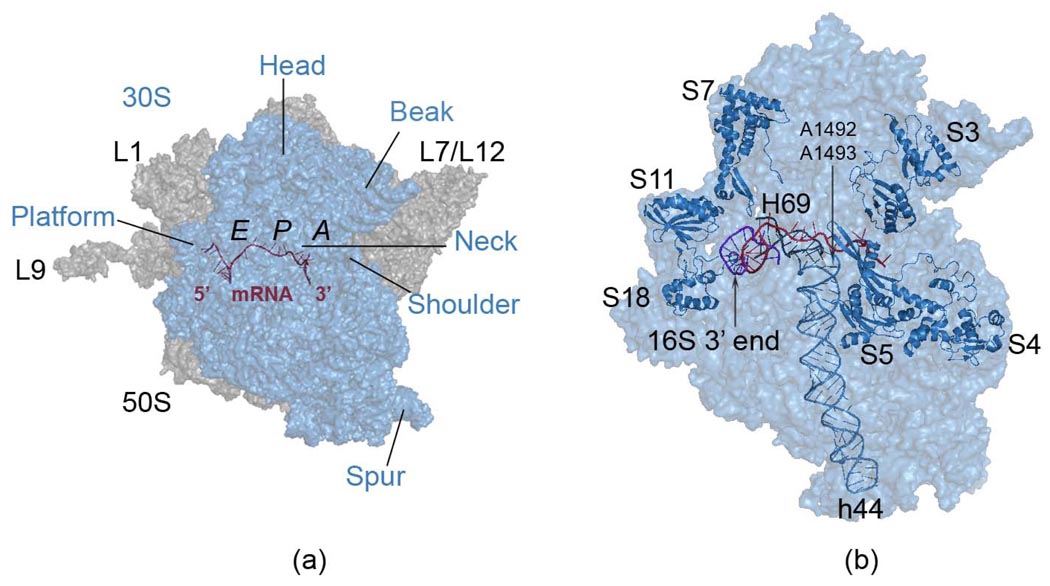
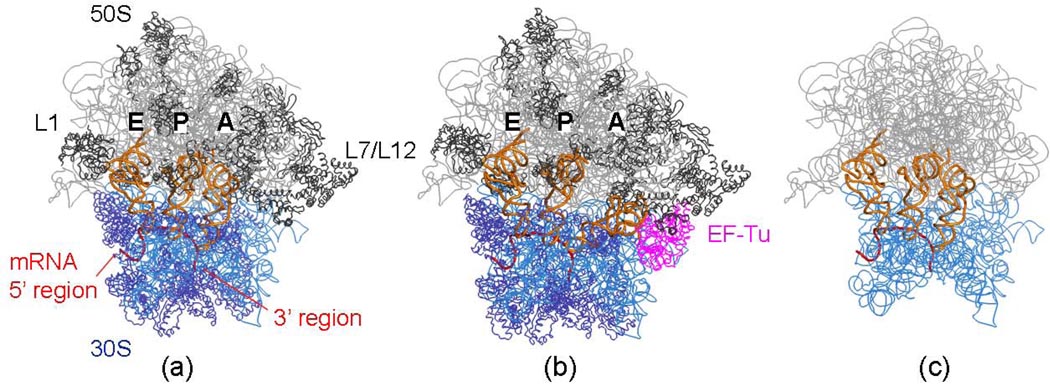
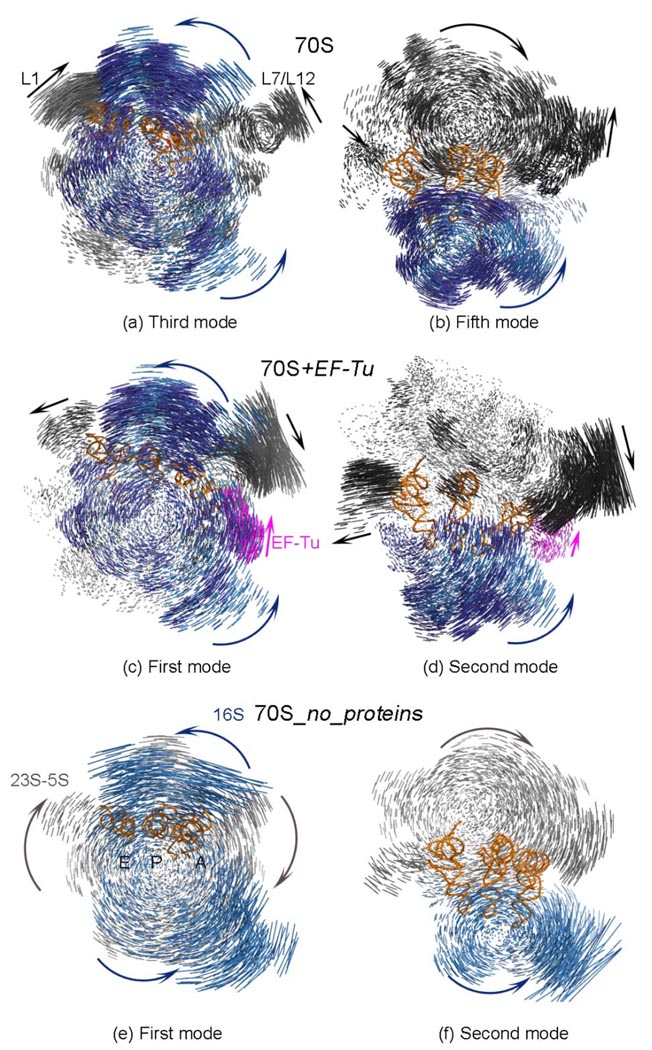
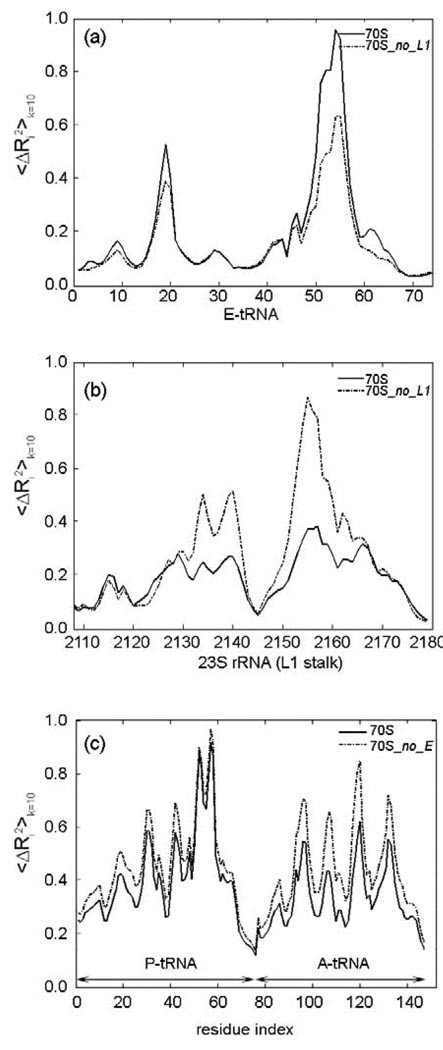
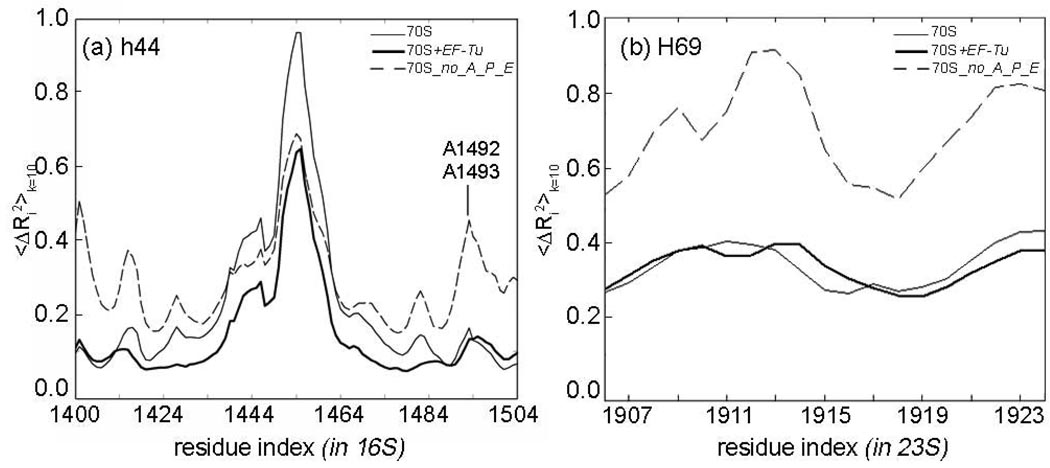
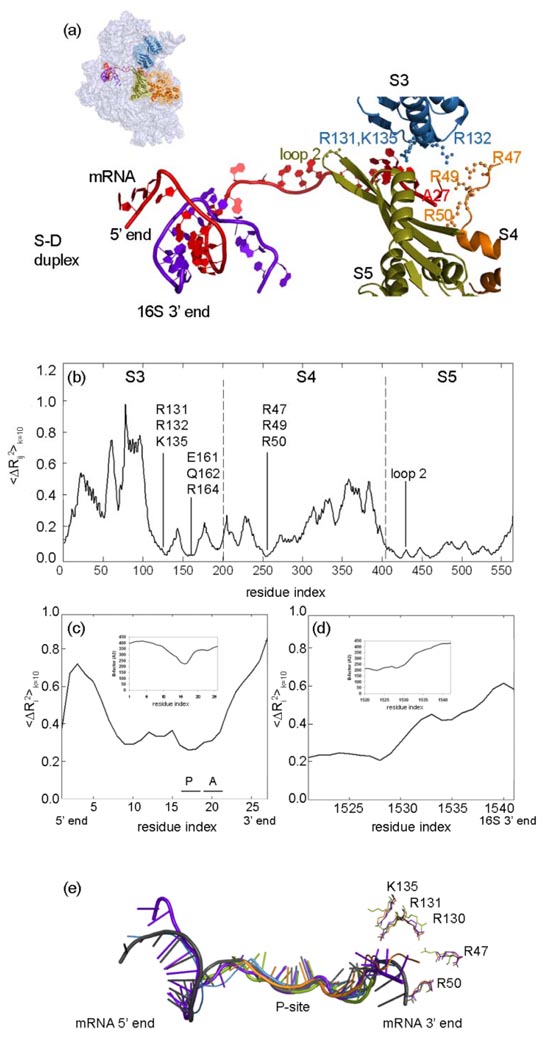
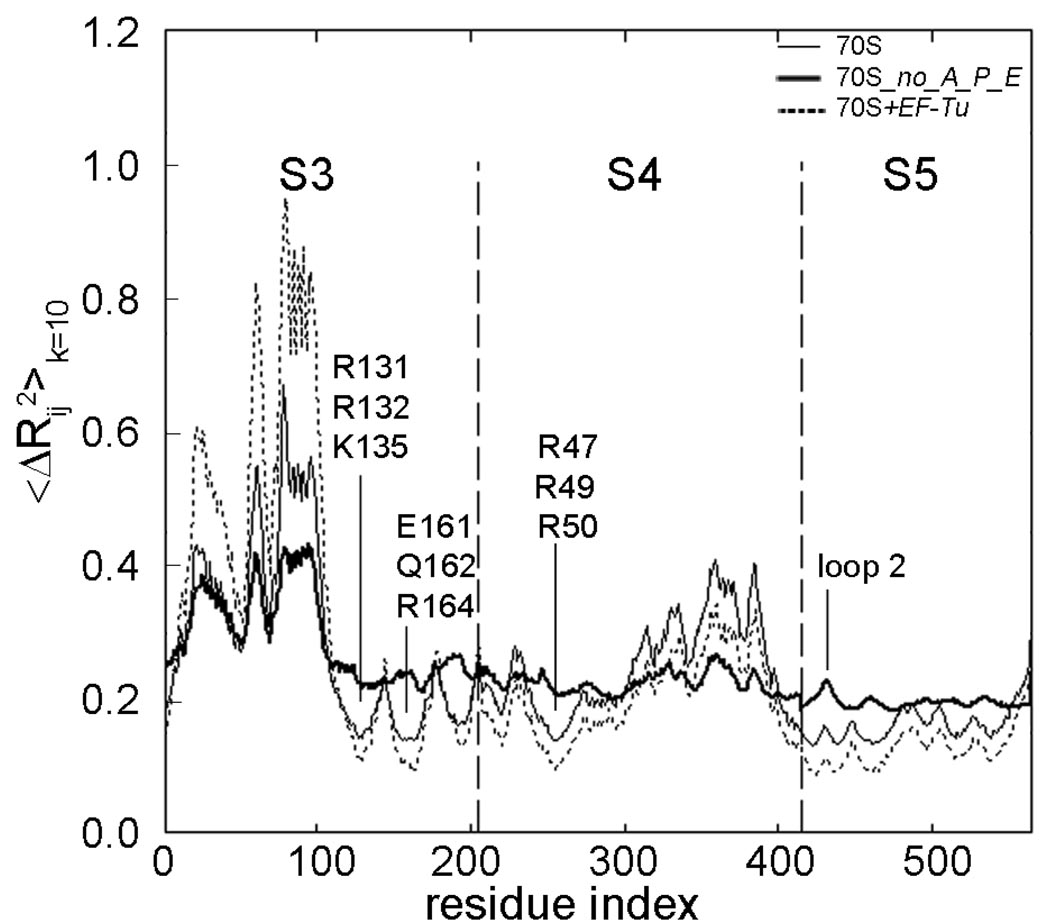
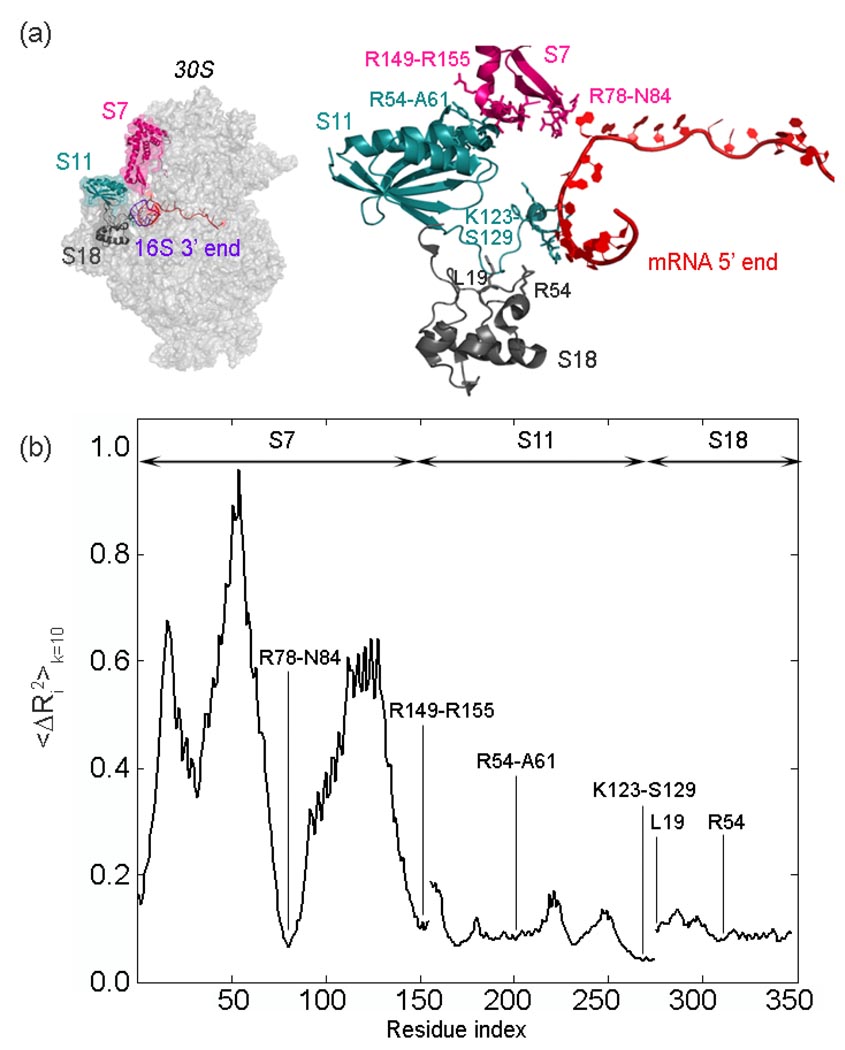
The protein-synthesizing ribosome undergoes large motions to effect the translocation of tRNAs and mRNA; here, the domain motions of this system are explored with a coarse-grained elastic network model using normal mode analysis. Crystal structures are used to construct various model systems of the 70S complex with/without tRNA, elongation factor Tu and the ribosomal proteins. Computed motions reveal the well-known ratchet-like rotational motion of the large subunits, as well as the head rotation of the small subunit and the high flexibility of the L1 and L7/L12 stalks, even in the absence of ribosomal proteins. This result indicates that these experimentally observed motions during translocation are inherently controlled by the ribosomal shape and only partially dependent upon GTP hydrolysis. Normal mode analysis further reveals the mobility of A- and P-tRNAs to increase in the absence of the E-tRNA. In addition, the dynamics of the E-tRNA is affected by the absence of the ribosomal protein L1. The mRNA in the entrance tunnel interacts directly with helicase proteins S3 and S4, which constrain the mRNA in a clamp-like fashion, as well as with protein S5, which likely orients the mRNA to ensure correct translation. The ribosomal proteins S7, S11 and S18 may also be involved in assuring translation fidelity by constraining the mRNA at the exit site of the channel. The mRNA also interacts with the 16S 3' end forming the Shine-Dalgarno complex at the initiation step; the 3' end may act as a 'hook' to reel in the mRNA to facilitate its exit.
Figures
References
-
- Agrawal RK, Penczek P, Grassucci RA, Li Y, Leith A, Nierhaus KH, Frank J. Direct visualization of A-, P-, and E-site transfer RNAs in the Escherichia coli ribosome. Science. 1996;271:1000–1002. - PubMed
-
- Yusupov MM, Yusupova GZ, Baucom A, Lieberman K, Earnest TN, Cate JH, Noller HF. Crystal structure of the ribosome at 5.5 A resolution. Science. 2001;292:883–896. - PubMed
-
- Nissen P, Hansen J, Ban N, Moore PB, Steitz TA. The structural basis of ribosome activity in peptide bond synthesis. Science. 2000;289:920–930. - PubMed
-
- Agrawal RK, Heagle AB, Penczek P, Grassucci RA, Frank J. EF-G-dependent GTP hydrolysis induces translocation accompanied by large conformational changes in the 70S ribosome. Nat. Struct. Biol. 1999;6:643–647. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources