

Review
doi: 10.1039/b717196j.
Epub 2008 Sep 9.
The chemistry and biochemistry of heme c: functional bases for covalent attachment
Affiliations
- PMID: 19030605
- PMCID: PMC2654777
- DOI: 10.1039/b717196j
Item in Clipboard
Review
The chemistry and biochemistry of heme c: functional bases for covalent attachment
Nat Prod Rep.
2008 Dec.
Abstract
A discussion of the literature concerning the synthesis, function, and activity of heme c-containing proteins is presented. Comparison of the properties of heme c, which is covalently bound to protein, is made to heme b, which is bound noncovalently. A question of interest is why nature uses biochemically expensive heme c in many proteins when its properties are expected to be similar to heme b. Considering the effects of covalent heme attachment on heme conformation and on the proximal histidine interaction with iron, it is proposed that heme attachment influences both heme reduction potential and ligand-iron interactions.
Figures
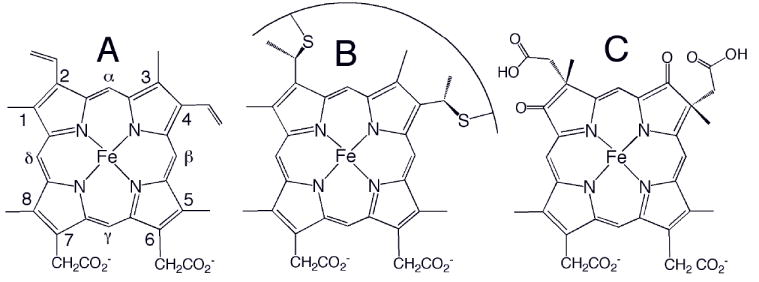
Chemical structures of (A) heme b, (B) heme c, and (C) heme d1. The Fisher numbering system for heme substituents is shown in (A). The curve in (B) represents a peptide segment; usually two residues separate the two Cys, and the His axial ligand follows the Cys attached to position 4. Heme o has a hydroxyfarnesyl group in place of vinyl 2 in (A), and heme a has a hydroxyfarnesyl group in place of vinyl 2 and a formyl group in place of methyl 8.
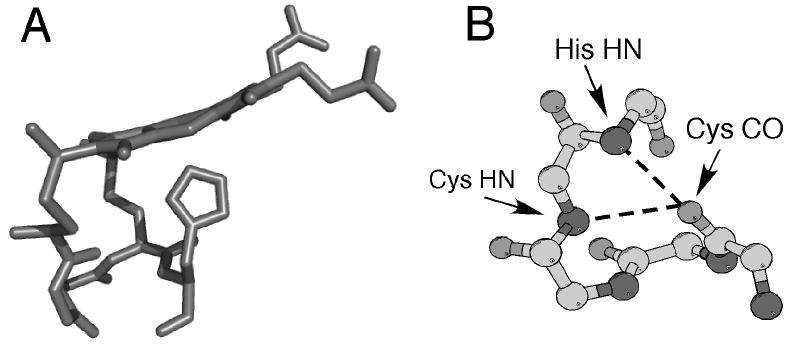
(A) Three-dimensional structure of heme c (side view) and the CXXCH (residues 14-18) pentapeptide from the structure of horse cytochrome c (PDB: 1HRC). The side chains of residues 15 and 16 are omitted for clarity. Note the nonplanar structure of the heme. (B) Ball-and-stick representation of backbone atoms of CXXCH motif. Hydrogen bonding interactions between the backbone HN of the second Cys and the His to the backbone CO of the first His are shown with dashed lines.
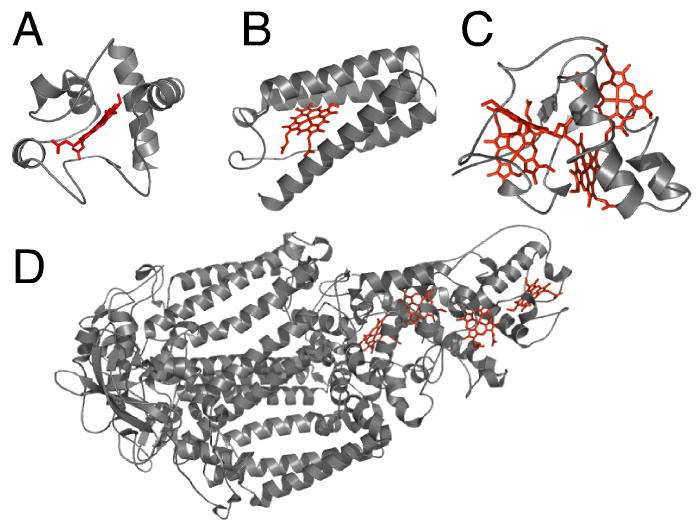
Three-dimensional structures of cytochromes c representative of each of the 4 structural classes. (A) Class 1: horse cytochrome c (PDB: 1HRC); (B) Class 2: Rhodopseudomonas capsulatus cytochrome c’ (1CPQ); (C) Class 3: Desulfovibrio vulgaris Hildenborough cytochrome c3 (1CTH); (D) Class 4: Photosynthetic reaction center from Rhodopseudomonas viridis (1PRC). In (D), cofactors other than the four hemes c are omitted for clarity. The polypeptides are shown in ribbon format and the hemes as sticks.
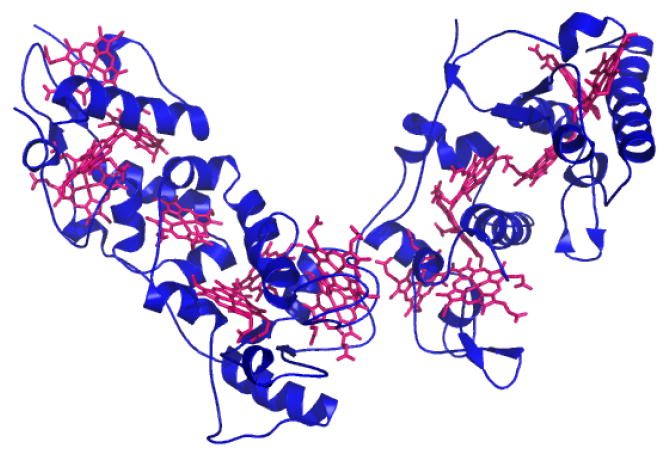
Three-dimensional structure of Desulfovibrio vulgaris Hildenborough HmcA (PDB: 1H29), a hexadecaheme cytochrome c, with hemes c shown in magenta.
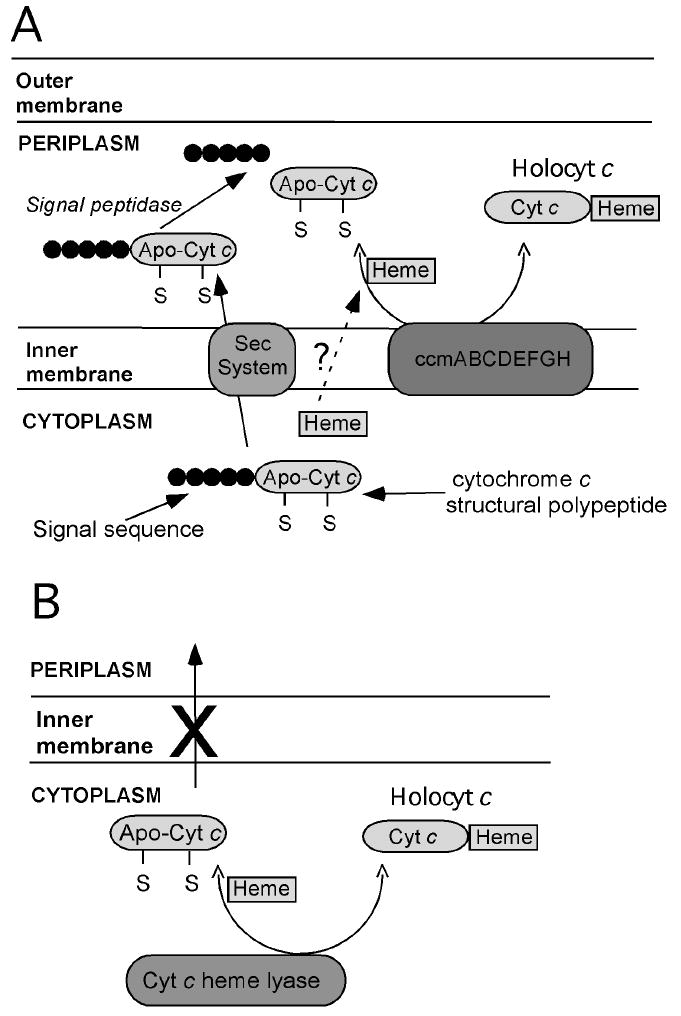
Schematic representation of systems for heterologous expression of cytochromes c in E. coli. (A) System based on E. coli Ccm machinery. A pre-apocytochrome c is expressed with an E. coli-compatible signal sequence directing translocation into the periplasm, where the Ccm apparatus assists with heme attachment. (B) System based on yeast CCHL. A cytochrome c structural gene is expressed along with yeast CCHL, which catalyzes heme attachment. As no signal sequence is used, heme attachment takes place in the cytoplasm.
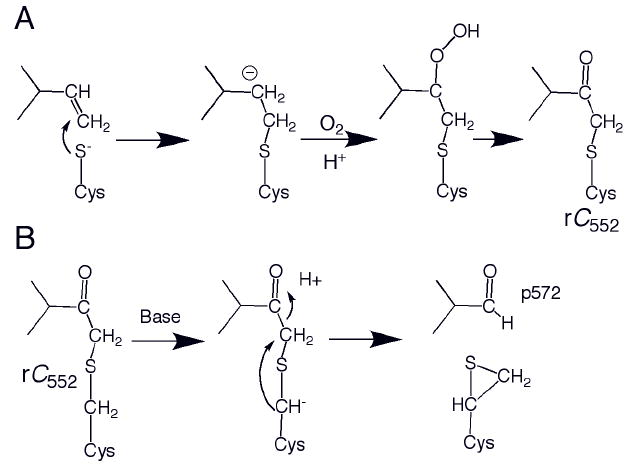
(A) Proposed nucleophilic mechanism for formation of Tt rC552. An analogous radial mechanism also is plausible. (B) Proposed mechanism for formation of p572 from Tt rC552. The proposed thiirane product shown has not been characterized.
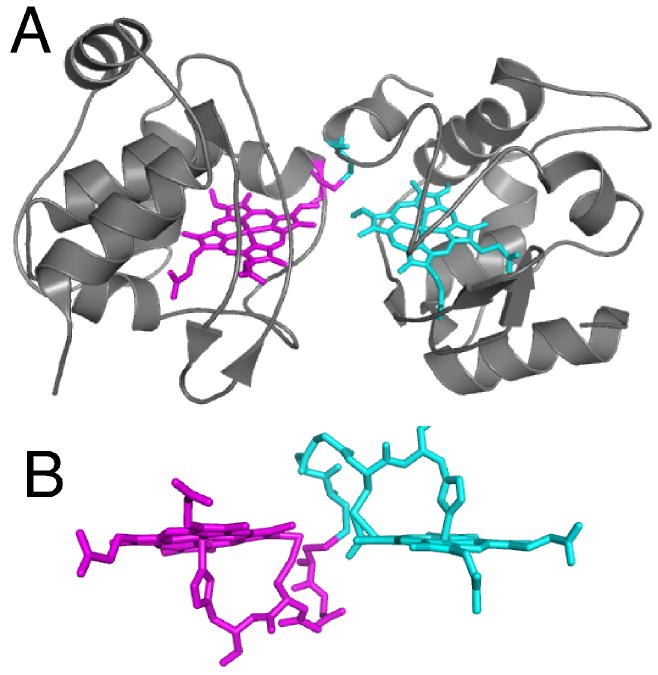
(A) Structure of T. thermophilus rC557 (PDB: 1FOC). Each heme is rotated about the α,γ-meso axis relative to the orientation in correctly matured cytochromes c, and each heme is attached via one thioether bond between 2-vinyl and Cys14. The Cys11 residues form an inter-protomer disulfide bond rather than binding to heme. (B) Detailed view of heme and residues 11-15 (CXXCH) in rC557. The atoms from each protomer are depicted in different colors. Note that the heme groups and CXXCH units from the two protomers are linked to each other via the interprotomer disulfide.
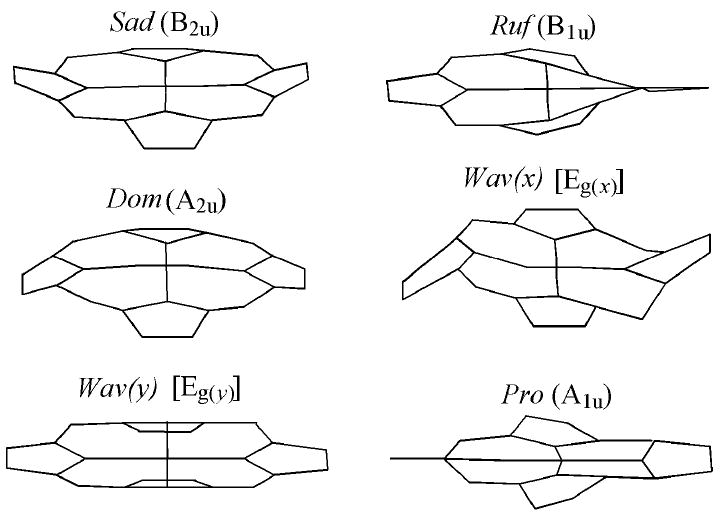
Out-of-plane modes representing nonplanar deformations of the porphyrin macrocycle, from work of Shelnutt and coworkers.
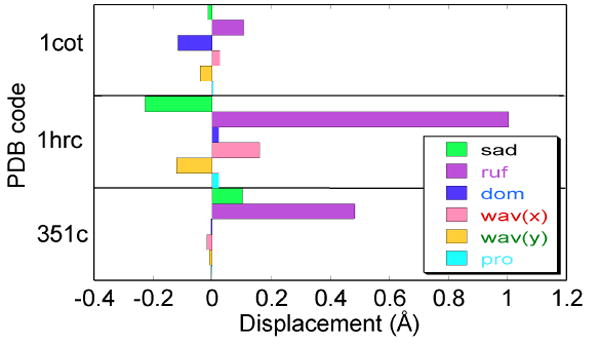
NSD results for horse heart cytochrome c (1HRC), P. aeruginosa cytochrome c551 (351C), and Paracoccus denitrificans cytochrome c2 (1COT). The primary mode of distortion of heme c usually is ruffling. However, the amount of ruffling varies substantially among cytochromes c, and the heme of cytochrome c2 is nearly planar.
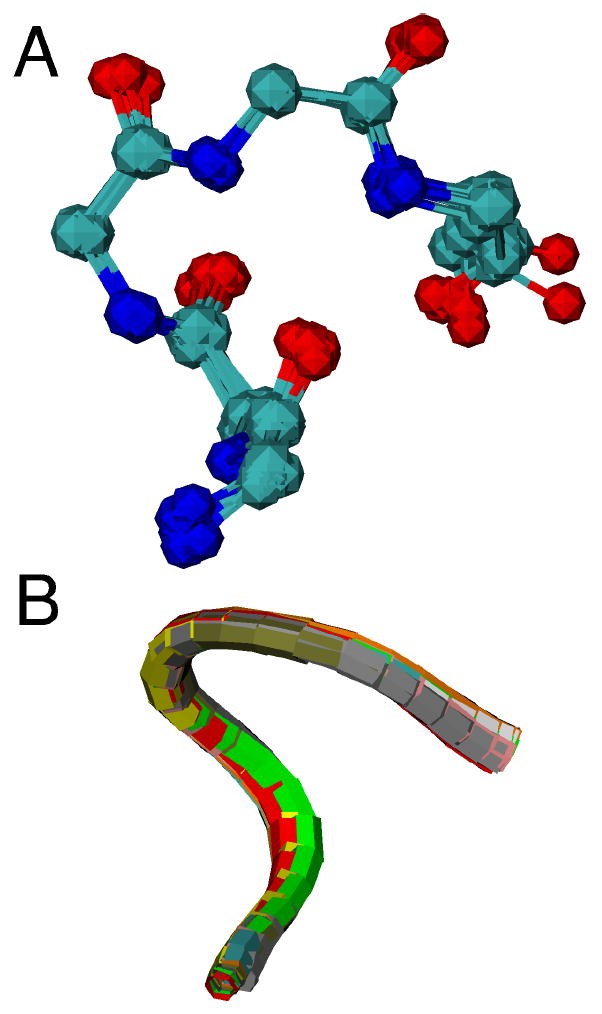
Overlay of CXXCH pentapeptide backbone for ten cytochromes c with diverse residues in the variable “XX” positions shown in (A) ball-and-stick and (B) ribbon format. Each protein is represented with a different color in (B). Proteins with crystal structures with 1.5-Å or better resolution were selected. The PDB accession codes and the identity of the variable “XX” resides in the structures are: 1B7V (IS), 1GU6 (AD), 1IQC (NS), 1YNR (MA), 1CTJ (AA), 2CTH (GD), 351C (VA), 1HRC (AQ), 1C2R (KT), 2YCC (LQ).
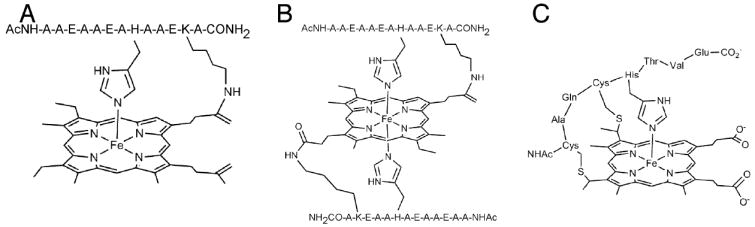
Examples of model systems utilized to study the effects of covalent attachment on the His-Fe interaction. (A, B) Peptide-sandwiched mesohemes (PSMs),, (C) Microperoxidase-8 (MP-8).
References
-
- Chapman SK, Daff S, Munro AW. Metal Sites in Proteins and Models. 1997:39–70.
-
- Schneider S, Marles-Wright J, Sharp KH, Paoli M. Nat Prod Rep. 2007;24:621–630. - PubMed
-
- Paoli M, Marles-Wright J, Smith A. DNA Cell Biol. 2002;21:271–280. - PubMed
-
- Stevens JM, Daltrop O, Allen JWA, Ferguson SJ. Acc Chem Res. 2004;37:999–1007. - PubMed
-
- Barker PD, Ferguson SJ. Structure. 1999;7:R281–R290. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources