

Functional insight into Maelstrom in the germline piRNA pathway: a unique domain homologous to the DnaQ-H 3'-5' exonuclease, its lineage-specific expansion/loss and evolutionarily active site switch
- PMID: 19032786
- PMCID: PMC2628886
- DOI: 10.1186/1745-6150-3-48
Functional insight into Maelstrom in the germline piRNA pathway: a unique domain homologous to the DnaQ-H 3'-5' exonuclease, its lineage-specific expansion/loss and evolutionarily active site switch
Abstract
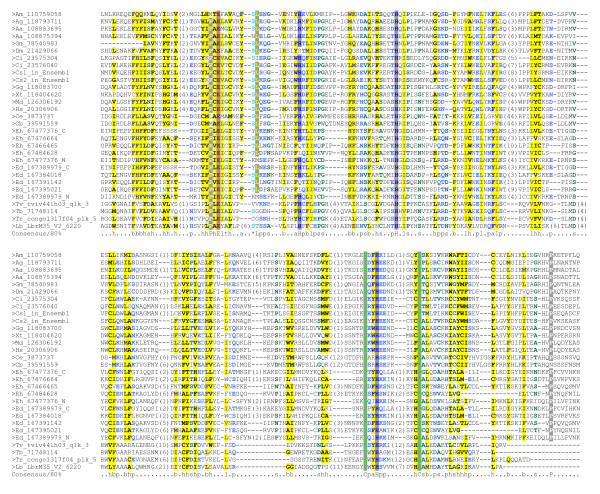
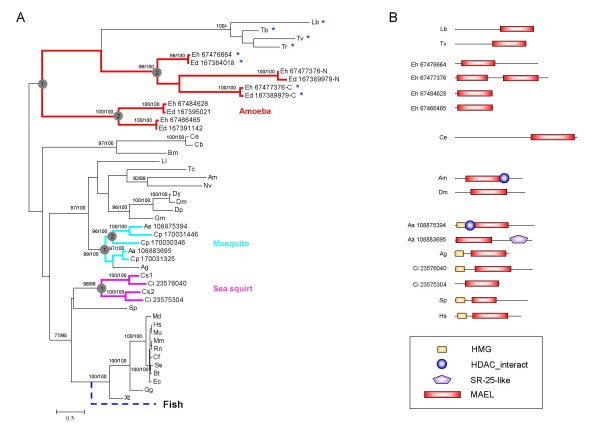
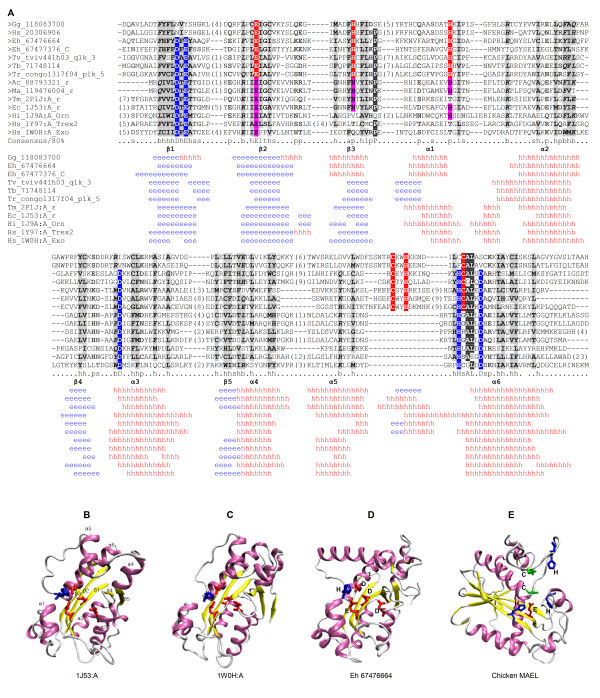
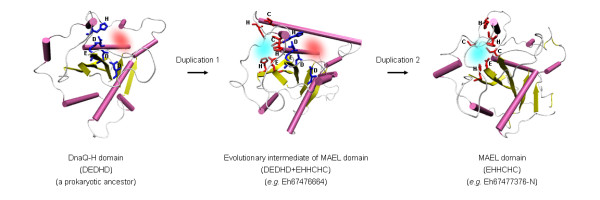
Maelstrom (MAEL) plays a crucial role in a recently-discovered piRNA pathway; however its specific function remains unknown. Here a novel MAEL-specific domain characterized by a set of conserved residues (Glu-His-His-Cys-His-Cys, EHHCHC) was identified in a broad range of species including vertebrates, sea squirts, insects, nematodes, and protists. It exhibits ancient lineage-specific expansions in several species, however, appears to be lost in all examined teleost fish species. Functional involvement of MAEL domains in DNA- and RNA-related processes was further revealed by its association with HMG, SR-25-like and HDAC_interact domains. A distant similarity to the DnaQ-H 3'-5' exonuclease family with the RNase H fold was discovered based on the evidence that all MAEL domains adopt the canonical RNase H fold; and several protist MAEL domains contain the conserved 3'-5' exonuclease active site residues (Asp-Glu-Asp-His-Asp, DEDHD). This evolutionary link together with structural examinations leads to a hypothesis that MAEL domains may have a potential nuclease activity or RNA-binding ability that may be implicated in piRNA biogenesis. The observed transition of two sets of characteristic residues between the ancestral DnaQ-H and the descendent MAEL domains may suggest a new mode for protein function evolution called "active site switch", in which the protist MAEL homologues are the likely evolutionary intermediates due to harboring the specific characteristics of both 3'-5' exonuclease and MAEL domains.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
