

MicroRNA-184 antagonizes microRNA-205 to maintain SHIP2 levels in epithelia
- PMID: 19033458
- PMCID: PMC2587229
- DOI: 10.1073/pnas.0803992105
MicroRNA-184 antagonizes microRNA-205 to maintain SHIP2 levels in epithelia
Abstract
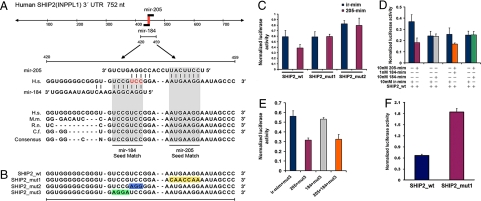
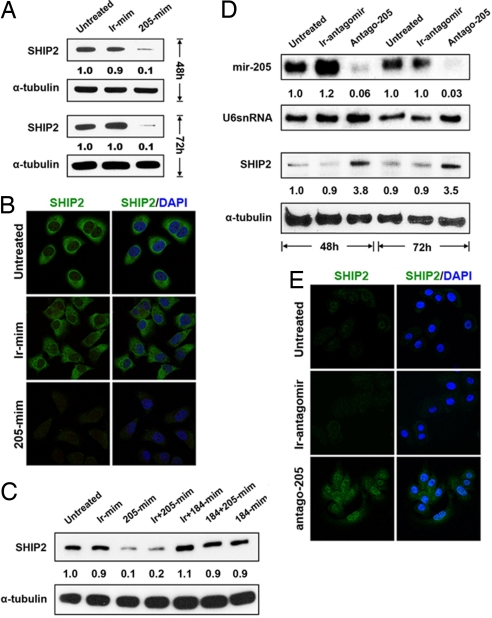
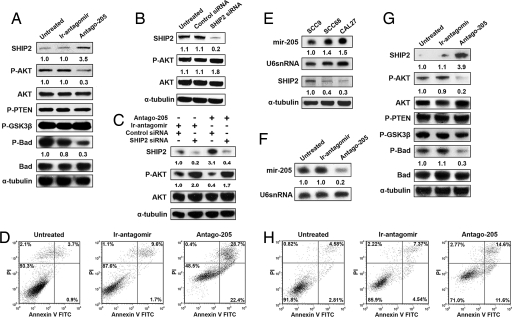
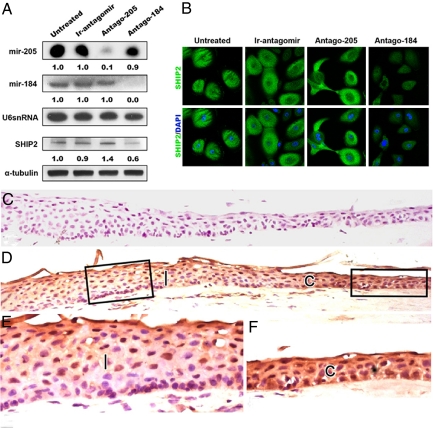
Despite their potential to regulate approximately one-third of the whole genome, relatively few microRNA (miRNA) targets have been experimentally validated, particularly in stratified squamous epithelia. Here we demonstrate not only that the lipid phosphatase SHIP2 is a target of miRNA-205 (miR-205) in epithelial cells, but, more importantly, that the corneal epithelial-specific miR-184 can interfere with the ability of miR-205 to suppress SHIP2 levels. This is the first example of a miRNA negatively regulating another to maintain levels of a target protein. Interfering with miR-205 function by using a synthetic antagomir, or by the ectopic expression of miR-184, leads to a coordinated damping of the Akt signaling pathway via SHIP2 induction. This was associated with a marked increase in keratinocyte apoptosis and cell death. Aggressive squamous cell carcinoma (SCC) cells exhibited elevated levels of miR-205. This was associated with a concomitant reduction in SHIP2 levels. Partial knockdown of endogenous miR-205 in SCCs markedly decreased phosphorylated Akt and phosphorylated BAD levels and increased apoptosis. We were able to increase SHIP2 levels in SCC cells after inhibition of miR-205. Therefore, miR-205 might have diagnostic value in determining the aggressivity of SCCs. Blockage of miR-205 activity with an antagomir or via ectopic expression of miR-184 could be novel therapeutic approaches for treating aggressive SCCs.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Ambros V. The functions of animal microRNAs. Nature. 2004;431:350–355. - PubMed
-
- Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. 1993;75:843–854. - PubMed
-
- Esau C, et al. MicroRNA-143 regulates adipocyte differentiation. J Biol Chem. 2004;279:52361–52365. - PubMed
-
- Hornstein E, et al. The microRNA miR-196 acts upstream of Hoxb8 and Shh in limb development. Nature. 2005;438:671–674. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
