

IRBIT coordinates epithelial fluid and HCO3- secretion by stimulating the transporters pNBC1 and CFTR in the murine pancreatic duct
- PMID: 19033647
- PMCID: PMC2613461
- DOI: 10.1172/JCI36983
IRBIT coordinates epithelial fluid and HCO3- secretion by stimulating the transporters pNBC1 and CFTR in the murine pancreatic duct
Abstract
Fluid and HCO3- secretion are vital functions of secretory epithelia. In most epithelia, this entails HCO3- entry at the basolateral membrane, mediated by the Na+-HCO3- cotransporter, pNBC1, and exit at the luminal membrane, mediated by a CFTR-SLC26 transporters complex. Here we report that the protein IRBIT (inositol-1,4,5-trisphosphate [IP3] receptors binding protein released with IP3), a previously identified activator of pNBC1, activates both the basolateral pNBC1 and the luminal CFTR to coordinate fluid and HCO3- secretion by the pancreatic duct. We used video microscopy and ion selective microelectrodes to measure fluid secretion and Cl- and HCO3- concentrations in cultured murine sealed intralobular pancreatic ducts. Short interference RNA-mediated knockdown of IRBIT markedly inhibited ductal pNBC1 and CFTR activities, luminal Cl- absorption and HCO3- secretion, and the associated fluid secretion. Single-channel measurements suggested that IRBIT regulated CFTR by reducing channel mean close time. Furthermore, expression of IRBIT constructs in HEK cells revealed that activation of pNBC1 required only the IRBIT PEST domain, while activation of CFTR required multiple IRBIT domains, suggesting that IRBIT activates these transporters by different mechanisms. These findings define IRBIT as a key coordinator of epithelial fluid and HCO3- secretion and may have implications to all CFTR-expressing epithelia and to cystic fibrosis.
Figures


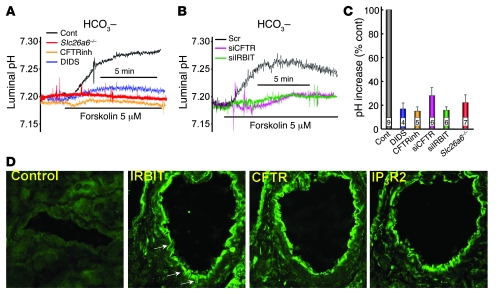
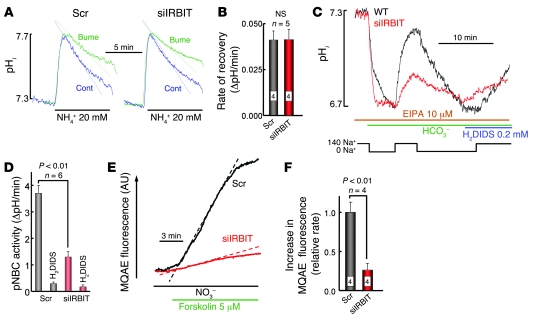


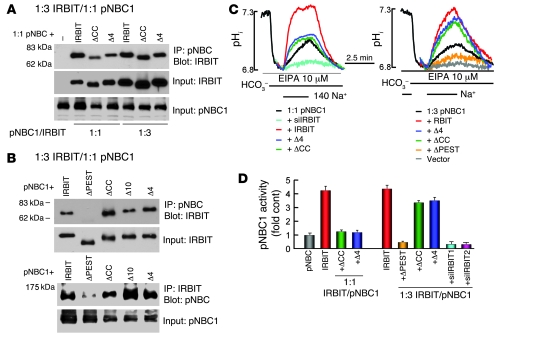
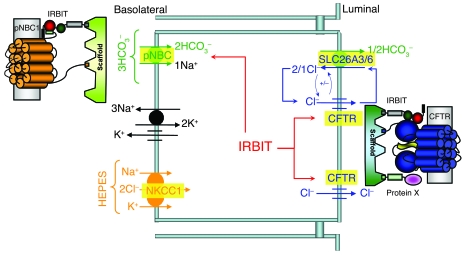
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
