

Vitamin D reduces the expression of collagen and key profibrotic factors by inducing an antifibrotic phenotype in mesenchymal multipotent cells
- PMID: 19036760
- PMCID: PMC3787314
- DOI: 10.1677/JOE-08-0241
Vitamin D reduces the expression of collagen and key profibrotic factors by inducing an antifibrotic phenotype in mesenchymal multipotent cells
Abstract
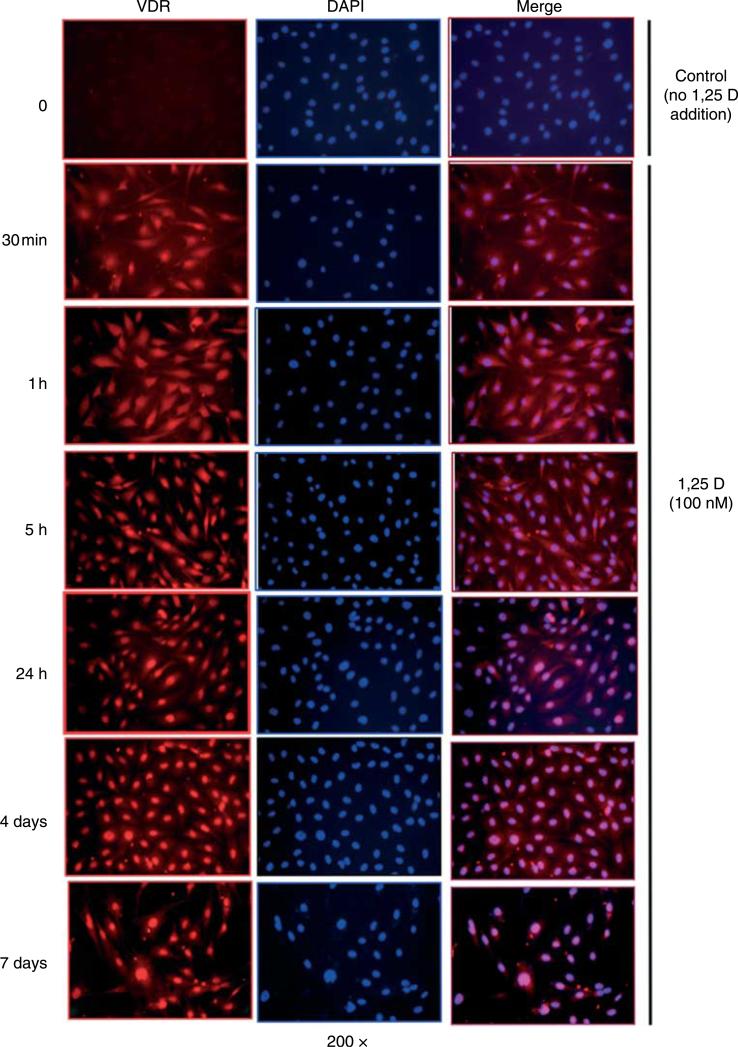
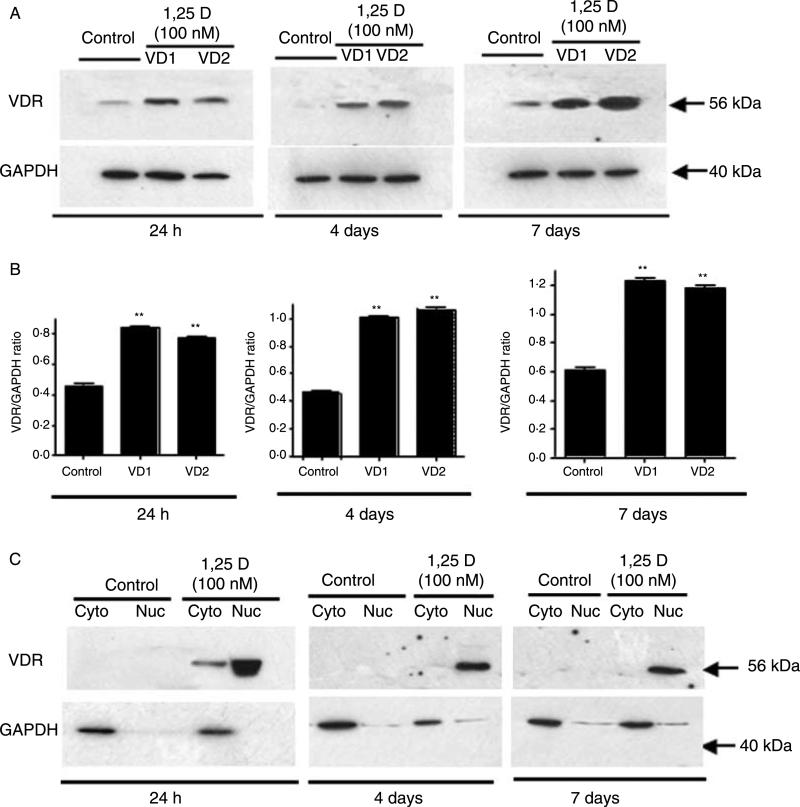
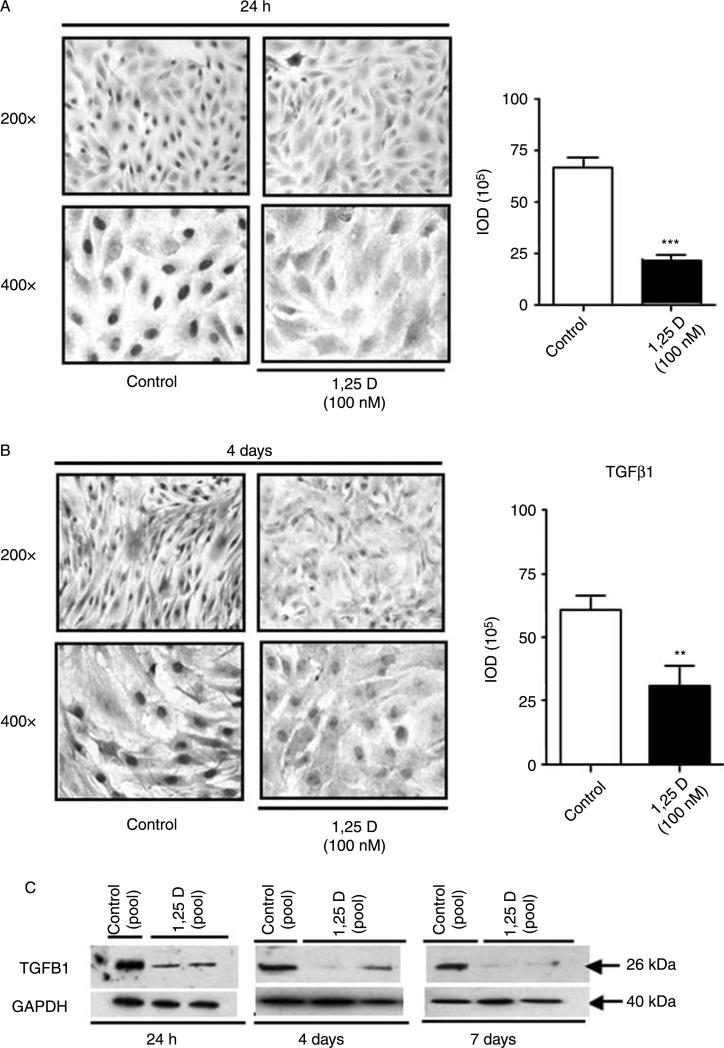
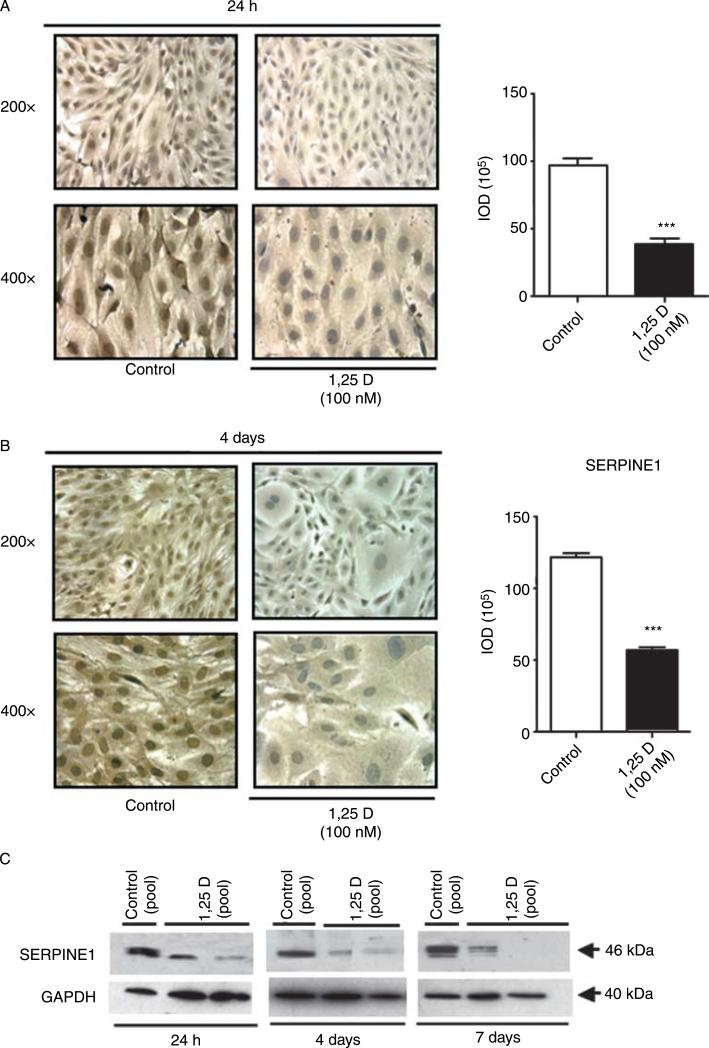
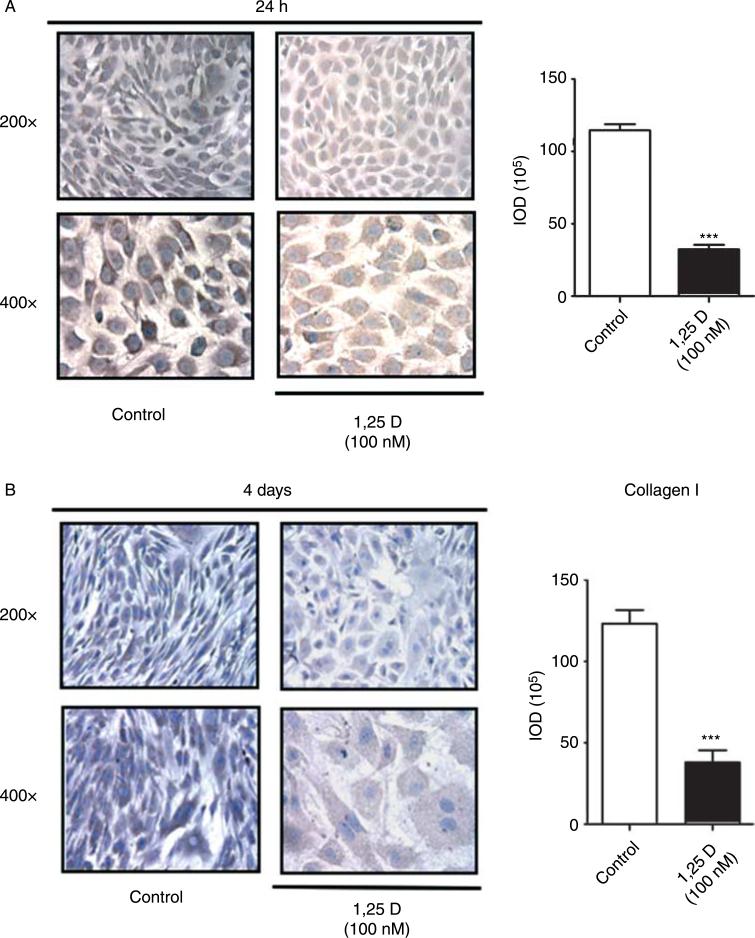
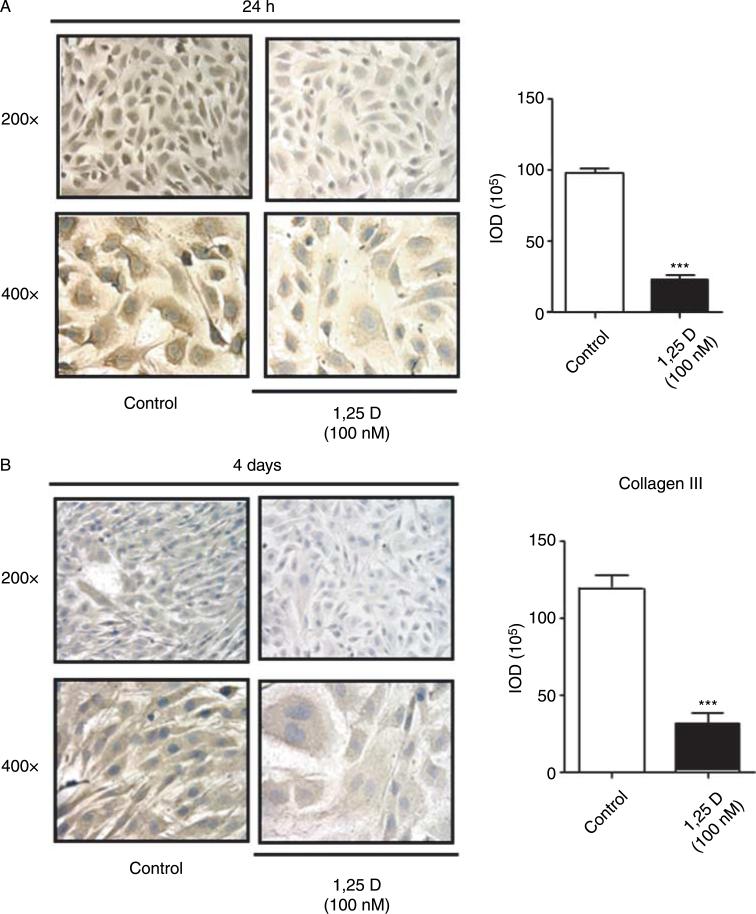
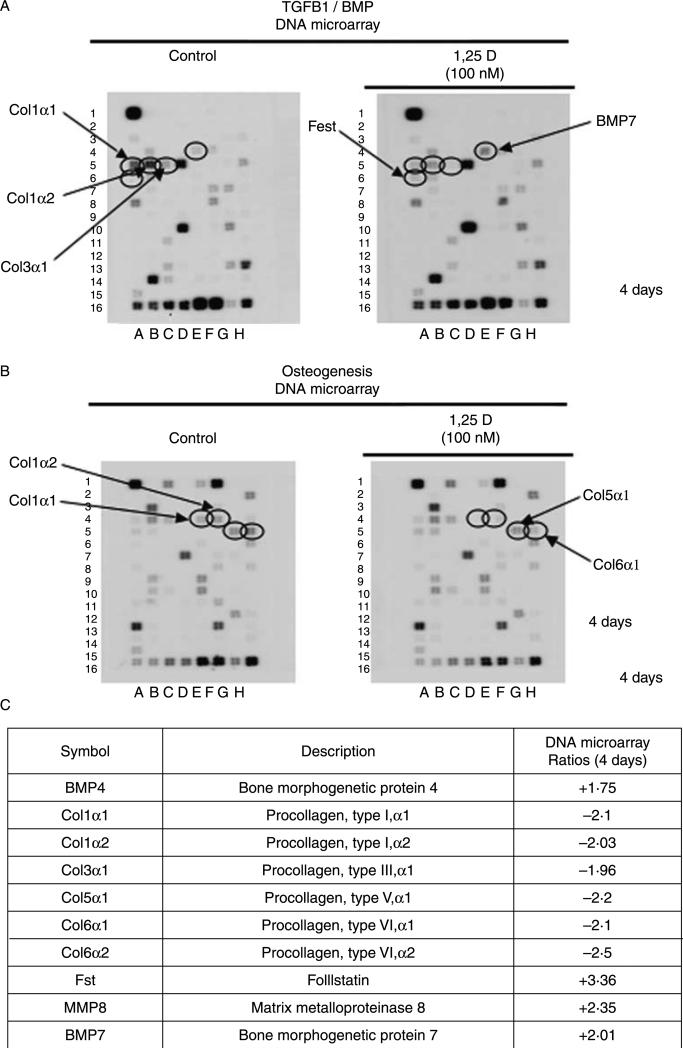
Hypovitaminosis D is an important public health problem. Serum 25-hydroxyvitamin D (25-OHD) is now recognized as an independent predictor for cardiovascular and related diseases (CVD) as well as other chronic medical conditions. However, the biologic pathways through which these effects are mediated remain poorly understood. We hypothesized that exposing mesenchymal multipotent cells (MMCs) to the active form of vitamin D would increase the expression of selected antifibrotic factors that in turn would ameliorate the progression of chronic diseases. MMCs were primed with 5'-azacytidine to induce a fibrotic phenotype and then treated with active vitamin D (1,25D) or ethanol <0.1% as vehicle in a time course manner (30 min, 1, 5, and 24 h, and for 4 and 7 days). The addition of 1,25D to MMCs promotes: a) increased expression and nuclear translocation of the vitamin D receptor; b) decreased expression of TGFB1 and plasminogen activator inhibitor (SERPINE1), two well-known profibrotic factors; c) decreased expression of collagen I, III and other collagens isoforms; and d) increased expression of several antifibrotic factors such as BMP7 a TGFB1 antagonist, MMP8 a collagen breakdown inducer and follistatin, an inhibitor of the profibrotic factor myostatin. In conclusion, the addition of 1,25D to differentiated MMCs displays a decreased profibrotic signaling pathway and gene expression, leading to decrease in collagen deposition. This study highlights key mechanistic pathways through which vitamin D decreases fibrosis, and provides a rationale for studies to test vitamin D supplementation as a preventive and/or early treatment strategy for CVD and related fibrotic disorders.
Figures

References
-
- Adams CD, Szumita PM, Baroletti SA, Lilly CM. Azacitidine-induced interstitial and alveolar fibrosis in a patient with myelodysplastic syndrome. Pharmacotherapy. 2005;25:765–768. - PubMed
-
- Amthor H, Nicholas G, McKinnell I, Kemp CF, Sharma M, Kambadur R, Patel K. Follistatin complexes myostatin and antagonises myostatin-mediated inhibition of myogenesis. Developmental Biology. 2004;270:19–30. - PubMed
-
- Aoki F, Kurabayashi M, Hasegawa Y, Kojima I. Attenuation of bleomycin-induced pulmonary fibrosis by follistatin. American Journal of Respiratory and Critical Care Medicine. 2005;172:713–720. - PubMed
-
- Artaza JN, Bhasin S, Mallidis C, Taylor W, Ma K, Gonzalez-Cadavid NF. Endogenous expression and localization of myostatin and its relation to myosin heavy chain distribution in C2C12 skeletal muscle cells. Journal of Cellular Physiology. 2002;190:170–179. - PubMed
-
- Artaza JN, Bhasin S, Magee TR, Reisz-Porszasz S, Shen R, Groome NP, Meerasahib MF, Gonzalez-Cadavid NF. Myostatin inhibits myogenesis and promotes adipogenesis in C3H 10T(1/2) mesenchymal multipotent cells. Endocrinology. 2005;146:3547–3557. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
