

MAVS dimer is a crucial signaling component of innate immunity and the target of hepatitis C virus NS3/4A protease
- PMID: 19036819
- PMCID: PMC2620913
- DOI: 10.1128/JVI.01659-08
MAVS dimer is a crucial signaling component of innate immunity and the target of hepatitis C virus NS3/4A protease
Abstract
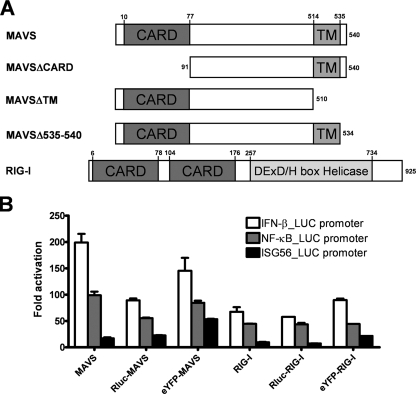
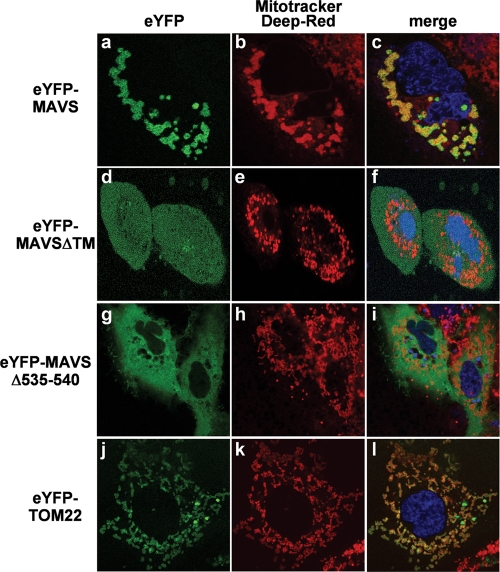
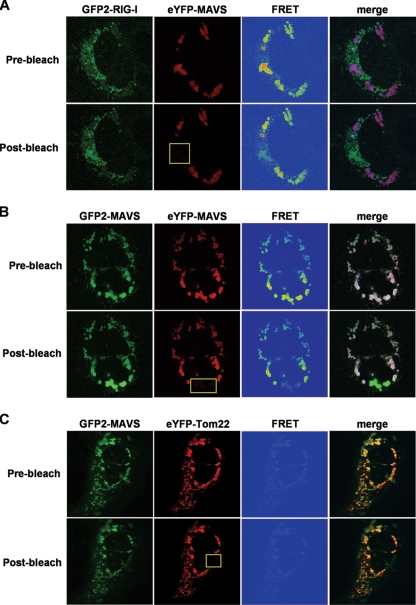
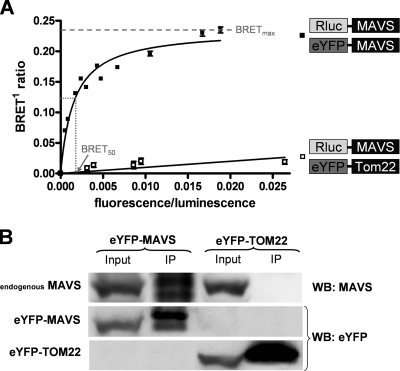
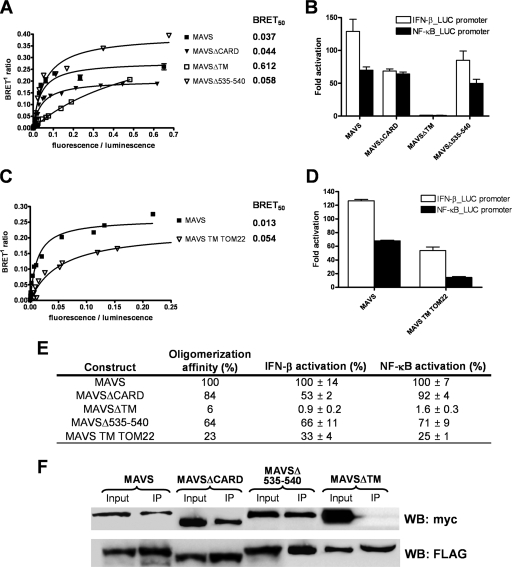
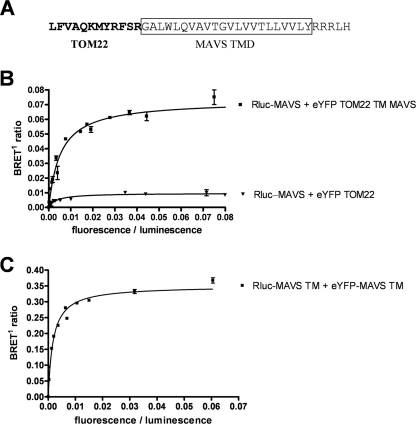
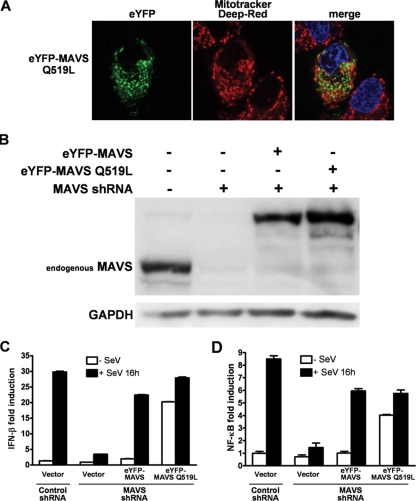
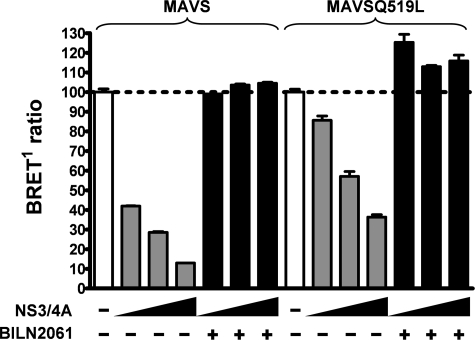
The mitochondrial antiviral signaling (MAVS) protein plays a central role in innate antiviral immunity. Upon recognition of a virus, intracellular receptors of the RIG-I-like helicase family interact with MAVS to trigger a signaling cascade. In this study, we investigate the requirement of the MAVS structure for enabling its signaling by structure-function analyses and resonance energy transfer approaches in live cells. We now report the essential role of the MAVS oligomer in signal transduction and map the transmembrane domain as the main determinant of dimerization. A combination of mutagenesis and computational methods identified a cluster of residues making favorable van der Waals interactions at the MAVS dimer interface. We also correlated the activation of IRF3 and NF-kappaB with MAVS oligomerization rather than its mitochondrial localization. Finally, we demonstrated that MAVS oligomerization is disrupted upon expression of HCV NS3/4A protease, suggesting a mechanism for the loss of antiviral signaling. Altogether, our data suggest that the MAVS oligomer is essential in the formation of a multiprotein membrane-associated signaling complex and enables downstream activation of IRF3 and NF-kappaB in antiviral innate immunity.
Figures

References
-
- Acehan, D., X. Jiang, D. G. Morgan, J. E. Heuser, X. Wang, and C. W. Akey. 2002. Three-dimensional structure of the apoptosome: implications for assembly, procaspase-9 binding, and activation. Mol. Cell 9423-432. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
