

Hormonal changes in relation to biomass partitioning and shoot growth impairment in salinized tomato (Solanum lycopersicum L.) plants
- PMID: 19036841
- PMCID: PMC2639025
- DOI: 10.1093/jxb/ern251
Hormonal changes in relation to biomass partitioning and shoot growth impairment in salinized tomato (Solanum lycopersicum L.) plants
Abstract
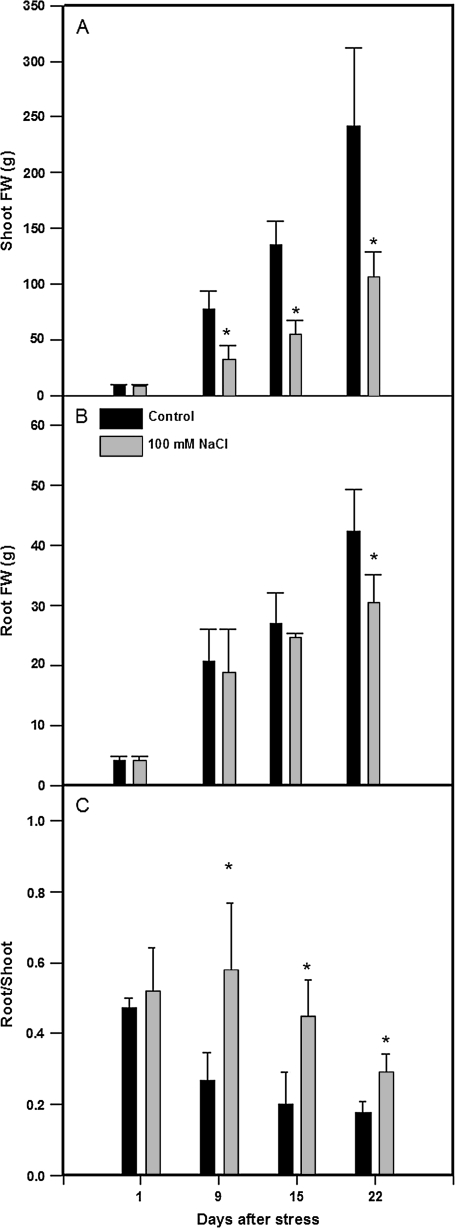
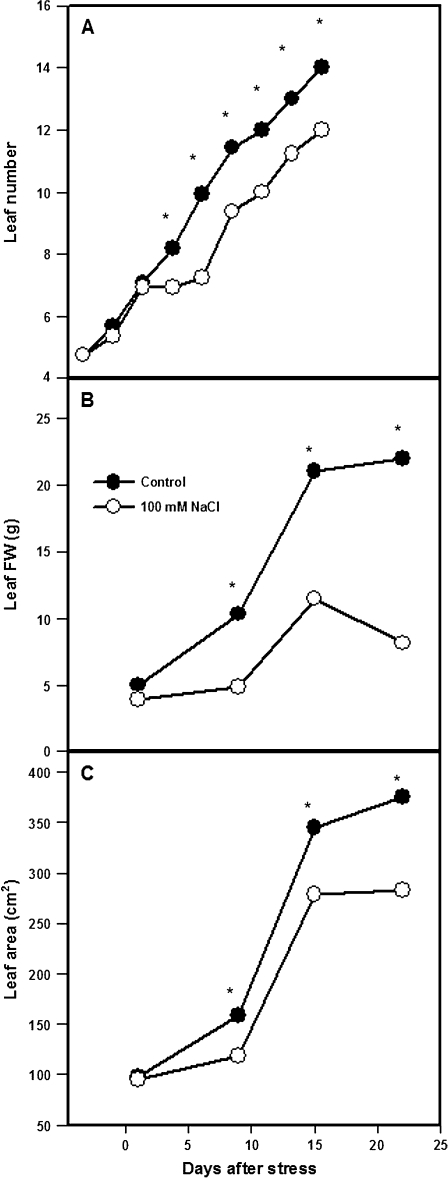
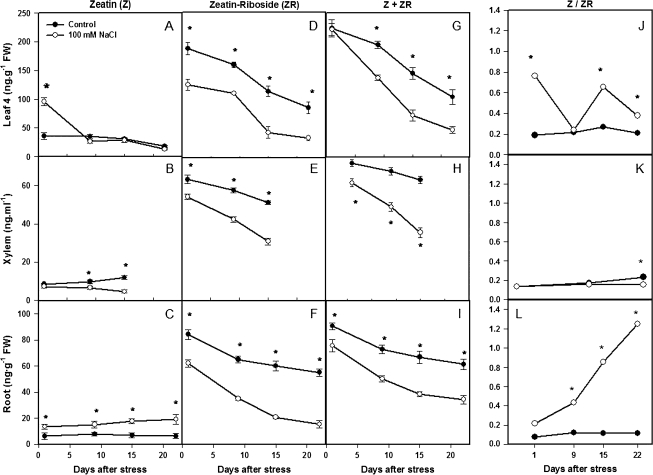
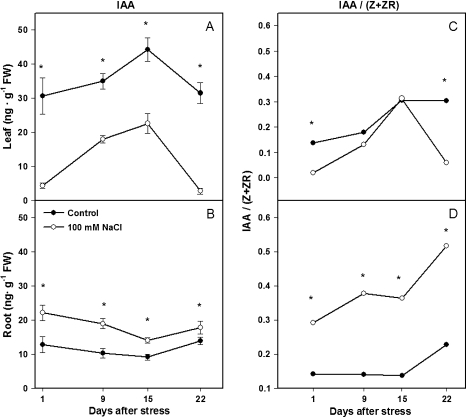
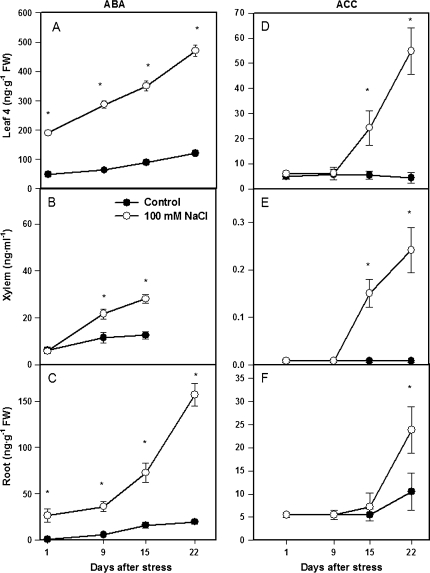
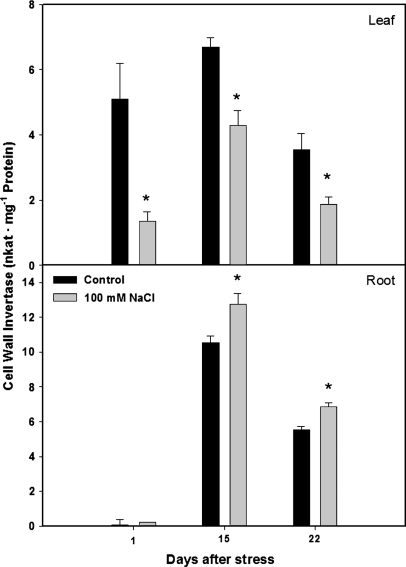
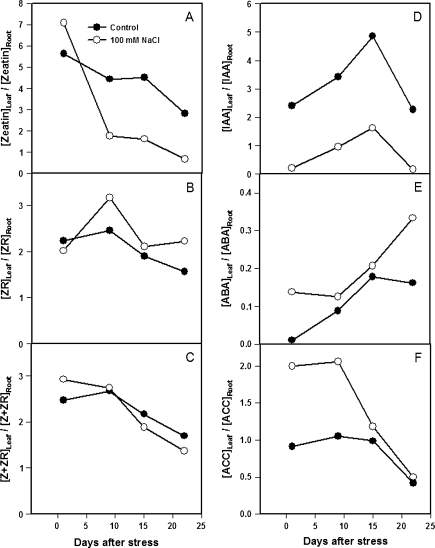
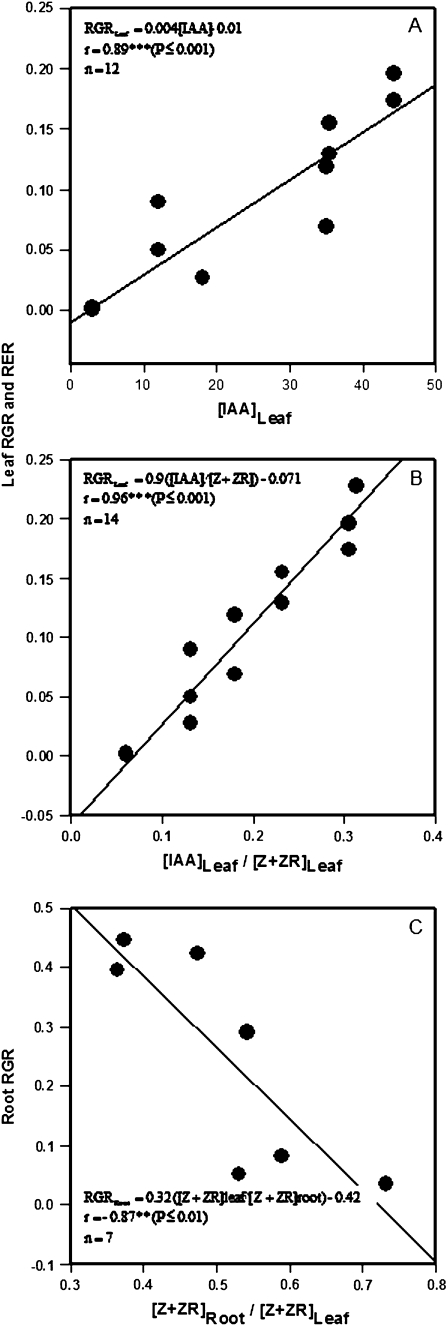
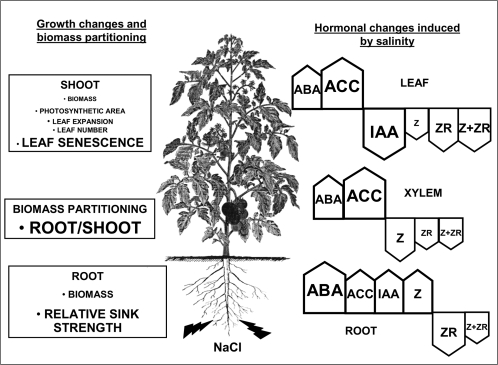
Following exposure to salinity, the root/shoot ratio is increased (an important adaptive response) due to the rapid inhibition of shoot growth (which limits plant productivity) while root growth is maintained. Both processes may be regulated by changes in plant hormone concentrations. Tomato plants (Solanum lycopersicum L. cv Moneymaker) were cultivated hydroponically for 3 weeks under high salinity (100 mM NaCl) and five major plant hormones (abscisic acid, ABA; the cytokinins zeatin, Z, and zeatin-riboside, ZR; the auxin indole-3-acetic acid, IAA; and the ethylene precursor 1-aminocyclopropane-1-carboxylic acid, ACC) were determined weekly in roots, xylem sap, and leaves. Salinity reduced shoot biomass by 50-60% and photosynthetic area by 20-25% both by decreasing leaf expansion and delaying leaf appearance, while root growth was less affected, thus increasing the root/shoot ratio. ABA and ACC concentrations strongly increased in roots, xylem sap, and leaves after 1 d (ABA) and 15 d (ACC) of salinization. By contrast, cytokinins and IAA were differentially affected in roots and shoots. Salinity dramatically decreased the Z+ZR content of the plant, and induced the conversion of ZR into Z, especially in the roots, which accounted for the relative increase of cytokinins in the roots compared to the leaf. IAA concentration was also strongly decreased in the leaves while it accumulated in the roots. Decreased cytokinin content and its transport from the root to the shoot were probably induced by the basipetal transport of auxin from the shoot to the root. The auxin/cytokinin ratio in the leaves and roots may explain both the salinity-induced decrease in shoot vigour (leaf growth and leaf number) and the shift in biomass allocation to the roots, in agreement with changes in the activity of the sink-related enzyme cell wall invertase.
Figures
References
-
- Balibrea ME, Dell'Amico J, Bolarín MC, Pérez-Alfocea F. Carbon partitioning and sucrose metabolism in tomato plants growing under salinity. Physiologia Plantarum. 2000;110:503–511.
-
- Balibrea ME, Parra M, Bolarín MC, Pérez-Alfocea F. Cytoplasmic sucrolytic activity controls tomato fruit growth under salinity. Australian Journal of Plant Physiology. 1999;26:561–568.
-
- Bangerth F. Response of cytokinin concentration in the xylem exudate of bean (Phaseolus vulgaris L.) plants to decapitation and auxin treatment and relationship to apical dominance. Planta. 1994;194:439–442.
-
- Beck EH. Regulation of the shoot/root ratio by cytokinins in Urtica dioica: opinion. Plant and Soil. 1996;185:3–12.
-
- Casas JL, Del Río JA, Serrano M, Acosta M. Comparison of 4 methods for the extraction and quantification of 1-aminocyclopropane-1-carboxylic acid (ACC) in tomato fruits. Revista de Agroquimica y Tecnología de Alimentos. 1989;29:191–198.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
