

Differential biochemical and cellular actions of Premarin estrogens: distinct pharmacology of bazedoxifene-conjugated estrogens combination
- PMID: 19036900
- PMCID: PMC5419325
- DOI: 10.1210/me.2008-0366
Differential biochemical and cellular actions of Premarin estrogens: distinct pharmacology of bazedoxifene-conjugated estrogens combination
Abstract
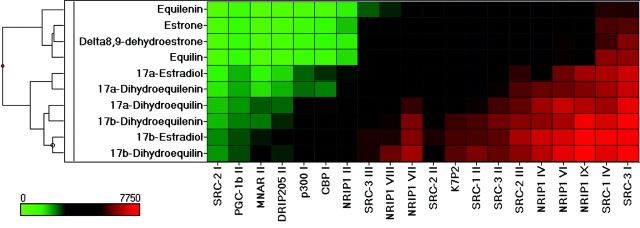
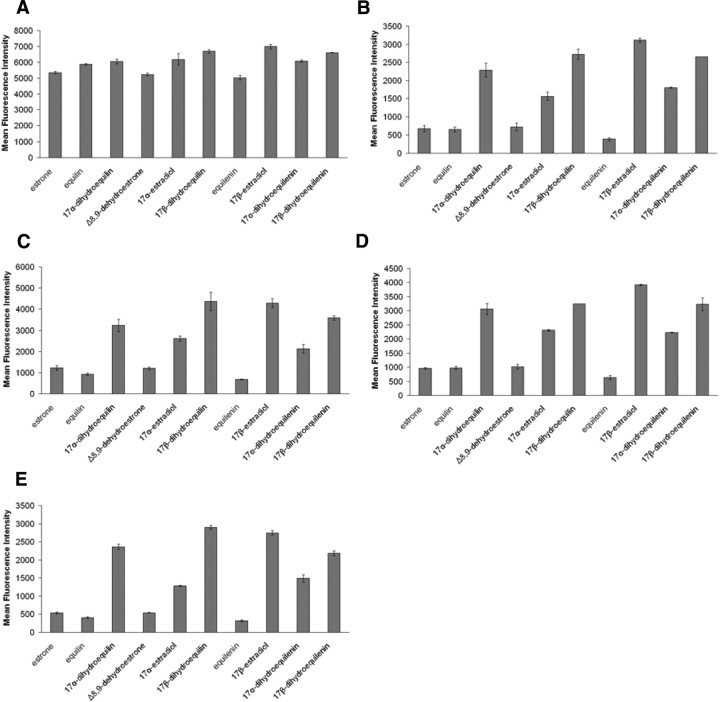
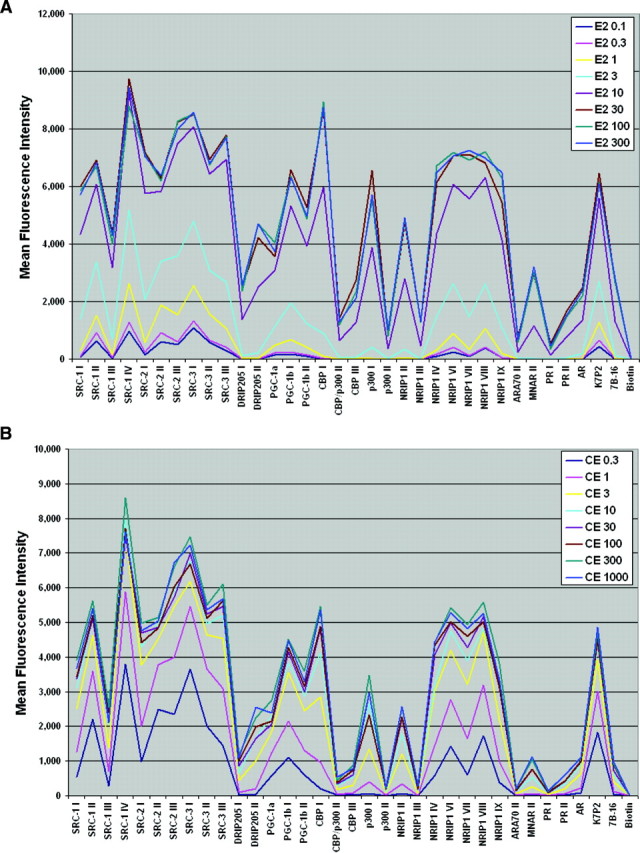
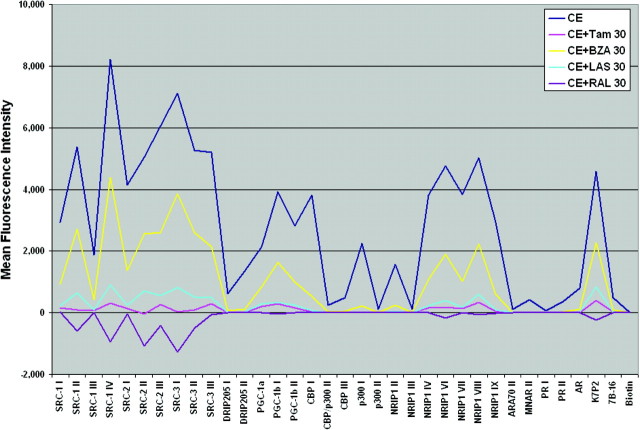
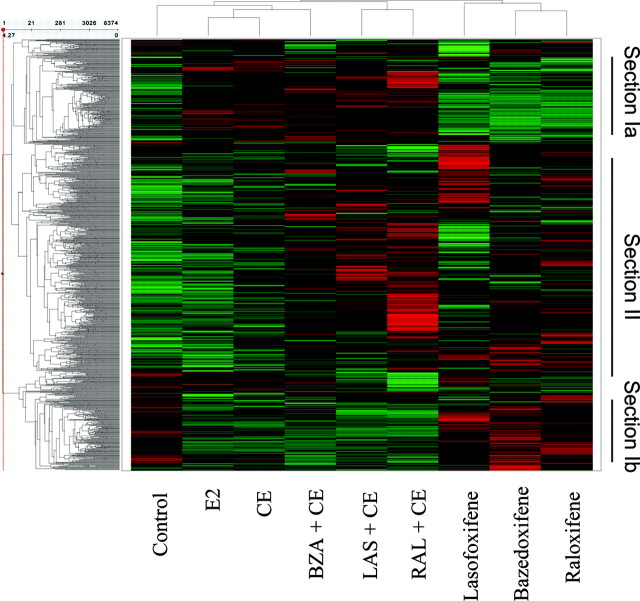
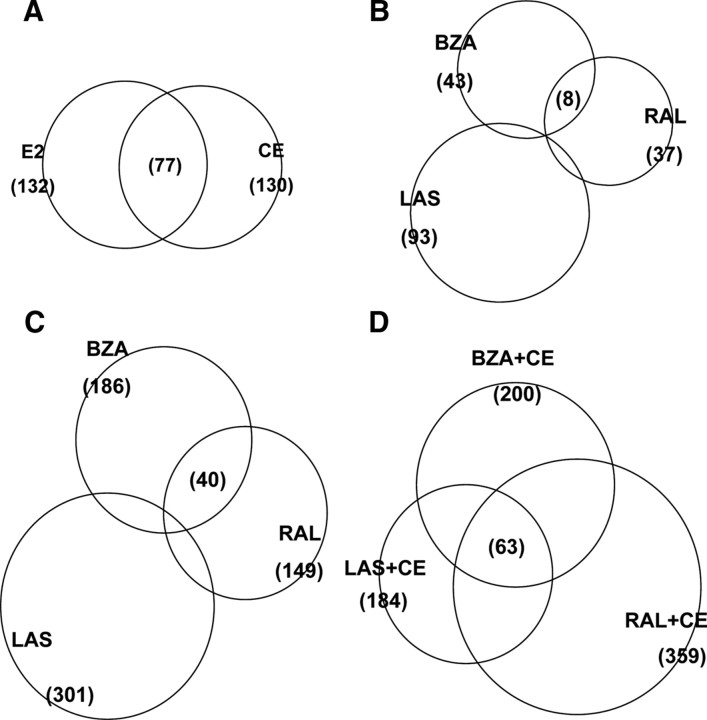
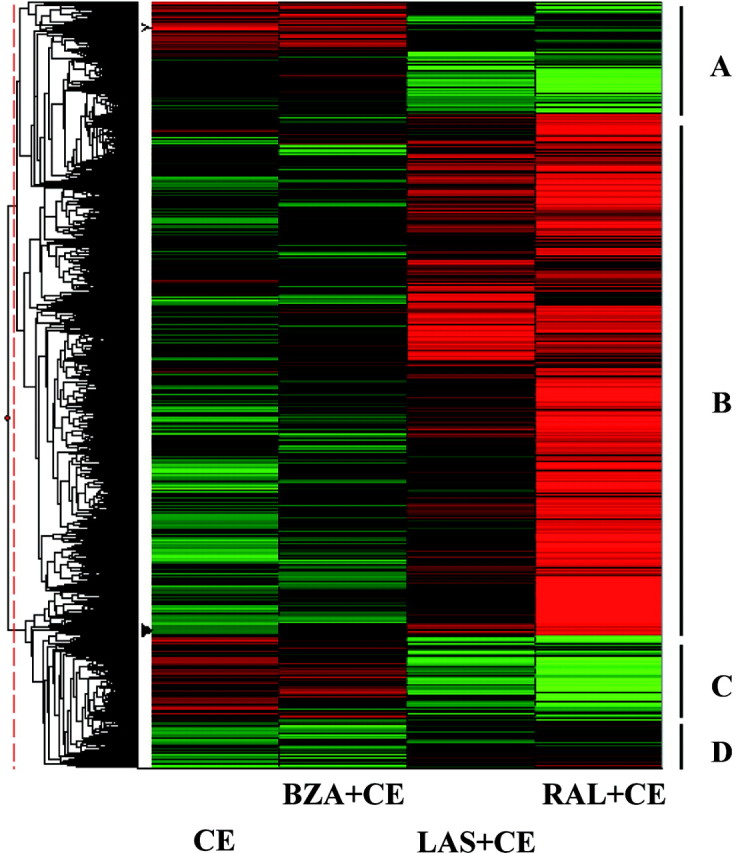
The use of estrogen-based therapies and the selective estrogen receptor (ER) modulator (SERM), raloxifene, which are approved for postmenopausal osteoporosis, is associated with side effects such as uterine/breast hyperproliferation, thromboembolism, and hot flashes. A combination of a new SERM, bazedoxifene (BZA), and Premarin (conjugated estrogens; CE) is under investigation to mitigate the estrogen/SERM side effects with promising results in Phase III clinical trials. To explore the mechanism of BZA/CE action, we investigated the recruitment of cofactor peptides to ERalpha by components of CE and a mixture containing the 10 major components of CE with or without three different SERMs. Here, we demonstrate differential recruitment of cofactor peptides to ERalpha by the individual CE components using a multiplex nuclear receptor-cofactor peptide interaction assay. We show that estrone and equilin are partial agonists in comparison with 17beta-estradiol in recruiting cofactor peptides to ERalpha. Further, CE was more potent than 17beta-estradiol in mediating ERalpha interaction with cofactor peptides. Interestingly, BZA was less potent than other SERMs in antagonizing the CE-mediated cofactor peptide recruitment to ERalpha. Finally, in accordance with these biochemical findings, 17beta-estradiol and CE, as well as SERM/CE combinations, showed differential gene regulation patterns in MCF-7 cells. In addition, BZA showed antagonism of a unique set of CE-regulated genes and did not down-regulate the expression of a number of CE-regulated genes, the expression of which was effectively antagonized by the other two SERMs. These results indicate that SERMs in combination with CE exhibit differential pharmacology, and therefore, combinations of other SERMs and estrogen preparations may not yield the same beneficial effects that are observed in clinic by pairing BZA with CE.
Figures
Similar articles
-
Gene expression profiling studies of three SERMs and their conjugated estrogen combinations in human breast cancer cells: insights into the unique antagonistic effects of bazedoxifene on conjugated estrogens.J Steroid Biochem Mol Biol. 2010 Jan;118(1-2):117-24. doi: 10.1016/j.jsbmb.2009.11.003. Epub 2009 Nov 13. J Steroid Biochem Mol Biol. 2010. PMID: 19914376
-
Differential utilization of nuclear and extranuclear receptor signaling pathways in the actions of estrogens, SERMs, and a tissue-selective estrogen complex (TSEC).J Steroid Biochem Mol Biol. 2016 Apr;158:198-206. doi: 10.1016/j.jsbmb.2015.12.008. Epub 2015 Dec 12. J Steroid Biochem Mol Biol. 2016. PMID: 26689478
-
Tissue-selective estrogen complexes for postmenopausal women.Maturitas. 2013 Nov;76(3):213-20. doi: 10.1016/j.maturitas.2013.06.003. Epub 2013 Jul 11. Maturitas. 2013. PMID: 23849704 Review.
-
The pairing of a selective estrogen receptor modulator, bazedoxifene, with conjugated estrogens as a new paradigm for the treatment of menopausal symptoms and osteoporosis prevention.Endocrinology. 2008 Dec;149(12):6084-91. doi: 10.1210/en.2008-0817. Epub 2008 Aug 14. Endocrinology. 2008. PMID: 18703623
-
Perspective on prescribing conjugated estrogens/bazedoxifene for estrogen-deficiency symptoms of menopause: a practical guide.Maturitas. 2015 Apr;80(4):435-40. doi: 10.1016/j.maturitas.2015.01.003. Epub 2015 Jan 24. Maturitas. 2015. PMID: 25684082 Review.
Cited by
-
Selective estrogen receptor modulators: tissue specificity and clinical utility.Clin Interv Aging. 2014 Aug 28;9:1437-52. doi: 10.2147/CIA.S66690. eCollection 2014. Clin Interv Aging. 2014. PMID: 25210448 Free PMC article. Review.
-
The evolution of selective estrogen receptor modulators in osteoporosis therapy.Climacteric. 2012 Dec;15(6):513-23. doi: 10.3109/13697137.2012.688079. Epub 2012 Aug 1. Climacteric. 2012. PMID: 22853318 Free PMC article. Review.
-
Tissue selective estrogen complexes (TSECs) differentially modulate markers of proliferation and differentiation in endometrial cells.Reprod Sci. 2013 Feb;20(2):129-37. doi: 10.1177/1933719112463251. Epub 2012 Nov 20. Reprod Sci. 2013. PMID: 23171676 Free PMC article.
-
Estrogens Promote Misfolded Proinsulin Degradation to Protect Insulin Production and Delay Diabetes.Cell Rep. 2018 Jul 3;24(1):181-196. doi: 10.1016/j.celrep.2018.06.019. Cell Rep. 2018. PMID: 29972779 Free PMC article.
-
Preventing osteoporosis with a tissue selective estrogen complex (TSEC) containing bazedoxifene/conjugated estrogens (BZA/CE).Osteoporos Int. 2011 Feb;22(2):447-51. doi: 10.1007/s00198-010-1440-x. Epub 2010 Nov 11. Osteoporos Int. 2011. PMID: 21069294 Review.
References
-
- Nelson HD2008. Menopause. Lancet 371:760–770 - PubMed
-
- Cooper C, O'Neill T, Silman A1993. The epidemiology of vertebral fractures. European Vertebral Osteoporosis Study Group. Bone 14(Suppl 1):S89–S97 - PubMed
-
- Melton LI1997. Epidemiology of spinal osteoporosis. Spine 22:2S–11S - PubMed
-
- Melton LJ, Kan SH, Frye MA, Wahner HW, O'Fallon WM, Riggs BL1989. Epidemiology of vertebral fractures in women. Am J Epidemiol 129:1000–1011 - PubMed
-
- Wasnich RD1996. Vertebral fracture epidemiology. Bone 18(Suppl 3):179S–S183 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
