

Chromatin associated sense and antisense noncoding RNAs are transcribed from the var gene family of virulence genes of the malaria parasite Plasmodium falciparum
- PMID: 19037012
- PMCID: PMC2612763
- DOI: 10.1261/rna.1080109
Chromatin associated sense and antisense noncoding RNAs are transcribed from the var gene family of virulence genes of the malaria parasite Plasmodium falciparum
Abstract
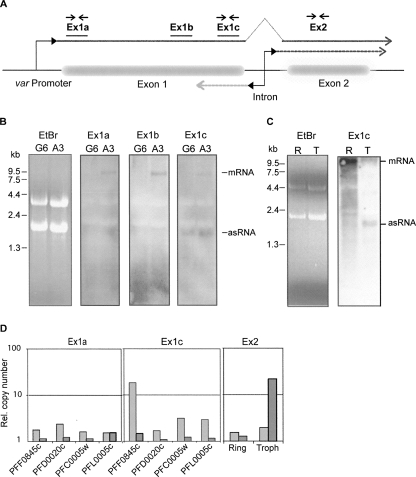
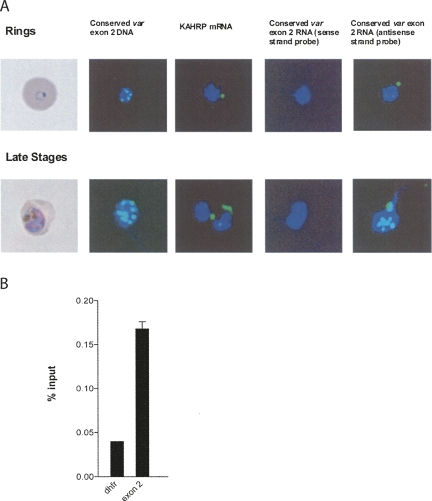
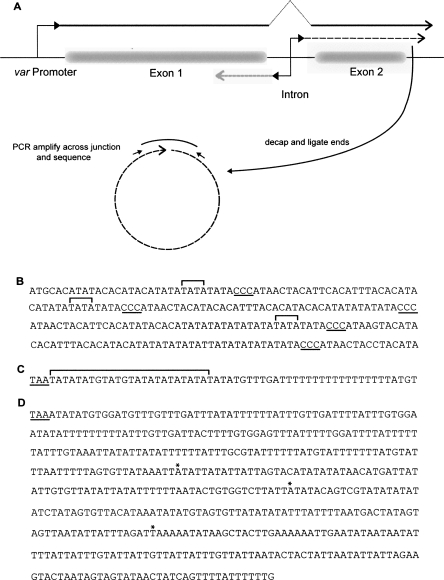
Antigenic variation by the malaria parasite Plasmodium falciparum results from switches in expression between members of the multicopy var gene family. These genes encode the variant surface protein PfEMP-1, the primary determinant of the antigenic and cytoadherent properties of infected erythrocytes. Only a single var gene is expressed at a time while the remaining members of the family remain transcriptionally silent. How mutually exclusive var gene expression is regulated is poorly understood; however, it is generally thought to involve alterations in chromatin assembly and modification, resulting in a type of cellular memory. Recently, several aspects of the chromatin structure surrounding var genes have been described, in particular the histone modifications associated with the active and silent states of the genes as well as their subnuclear localization. Here, we demonstrate that this chromatin structure also includes the incorporation of long sense and antisense noncoding RNAs. These sterile transcripts initiate from a bidirectional promoter located within a conserved intron found in all var genes that was previously implicated in var gene silencing. Mapping of the 5' and 3' ends of the sterile transcripts indicates that they are nonpolyadenylated. RNA fluorescent in situ hybridization (RNA-FISH) analysis detects both the sense and antisense noncoding RNAs in distinct spots within the nucleus similar to the pattern described for the var genes themselves. Further, analysis by RNA chromatin immunoprecipitation (ChIP) indicates that the noncoding RNAs are physically associated with chromatin. These sterile transcripts therefore might act in a manner analogous to noncoding RNAs associated with silent, condensed chromatin found in other eukaryotic systems.
Figures


References
-
- Akhtar A. Dosage compensation: An intertwined world of RNA and chromatin remodelling. Curr. Opin. Genet. Dev. 2003;13:161–169. - PubMed
-
- Andersson S.G.E., Zomorodipour A., Andersson J.O., Sicheritz-Ponten T., Alsmark U.C.M., Podowski R.M., Naslund A.K., Eriksson A.S., Winkler H.H., Kurland C.G. The genome sequence of Rickettsia prowazekii and the origin of mitochondria. Nature. 1998;396:133–140. - PubMed
-
- Aravind L., Iyer L.M., Wellems T.E., Miller L.H. Plasmodium biology: Genomic gleanings. Cell. 2003;115:771–785. - PubMed
-
- Baruch D.I., Pasloske B.L., Singh H.B., Bi X., Ma X.C., Feldman M., Taraschi T.F., Howard R.J. Cloning the P. falciparum gene encoding PfEMP1, a malarial variant antigen and adherence receptor on the surface of parasitized human erythrocytes. Cell. 1995;82:77–87. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources