

Early stages of Golgi vesicle and tubule formation require diacylglycerol
- PMID: 19037109
- PMCID: PMC2633395
- DOI: 10.1091/mbc.e08-03-0256
Early stages of Golgi vesicle and tubule formation require diacylglycerol
Abstract
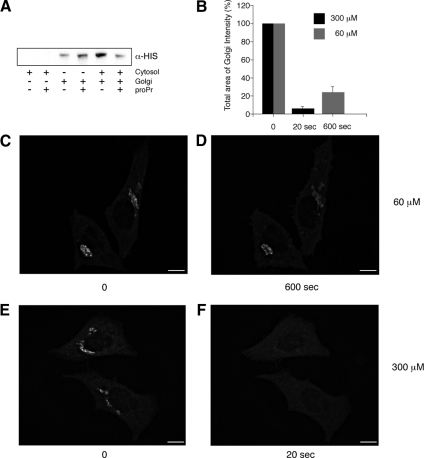
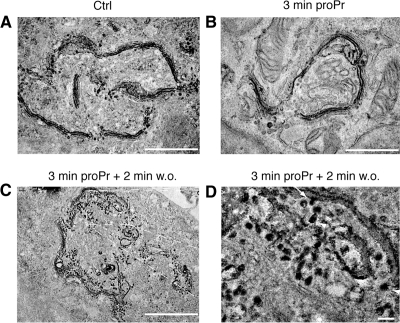
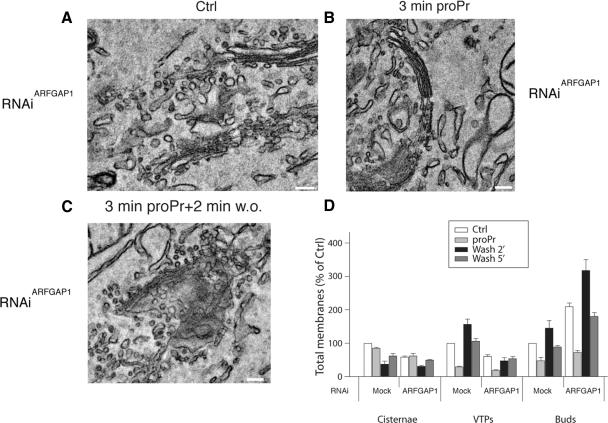
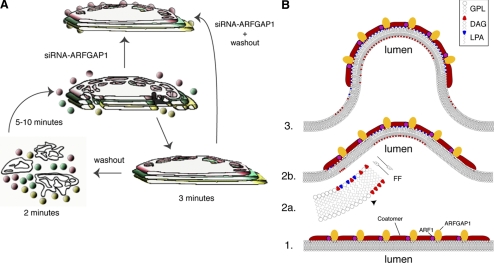
We have investigated the role for diacylglycerol (DAG) in membrane bud formation in the Golgi apparatus. Addition of propranolol to specifically inhibit phosphatidate phosphohydrolase (PAP), an enzyme responsible for converting phosphatidic acid into DAG, effectively prevents formation of membrane buds. The effect of PAP inhibition on Golgi membranes is rapid and occurs within 3 min. Removal of the PAP inhibitor then results in a rapid burst of buds, vesicles, and tubules that peaks within 2 min. The inability to form buds in the presence of propranolol does not appear to be correlated with a loss of ARFGAP1 from Golgi membranes, as knockdown of ARFGAP1 by RNA interference has little or no effect on actual bud formation. Rather, knockdown of ARFGAP1 results in an increase in membrane buds and a decrease of vesicles and tubules suggesting it functions in the late stages of scission. How DAG promotes bud formation is discussed.
Figures


References
-
- Antonny B., Bigay J., Casella J. F., Drin G., Mesmin B., Gounon P. Membrane curvature and the control of GTP hydrolysis in Arf1 during COPI vesicle formation. Biochem. Soc. Trans. 2005;33:619–622. - PubMed
-
- Antonny B., Huber I., Paris S., Chabre M., Cassel D. Activation of ADP-ribosylation factor 1 GTPase-activating protein by phosphatidylcholine-derived diacylglycerols. J. Biol. Chem. 1997;272:30848–30851. - PubMed
-
- Bai J., Pagano R. E. Measurement of spontaneous transfer and transbilayer movement of BODIPY-labeled lipids in lipid vesicles. Biochemistry. 1997;36:8840–8848. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
