

Gap junctions in the ovary of Drosophila melanogaster: localization of innexins 1, 2, 3 and 4 and evidence for intercellular communication via innexin-2 containing channels
- PMID: 19038051
- PMCID: PMC2631599
- DOI: 10.1186/1471-213X-8-111
Gap junctions in the ovary of Drosophila melanogaster: localization of innexins 1, 2, 3 and 4 and evidence for intercellular communication via innexin-2 containing channels
Abstract
Background: In the Drosophila ovary, germ-line and soma cells are interconnected via gap junctions. The main gap-junction proteins in invertebrates are members of the innexin family. In order to reveal the role that innexins play in cell-cell communication during oogenesis, we investigated the localization of innexins 1, 2, 3 and 4 using immunohistochemistry, and analyzed follicle development following channel blockade.
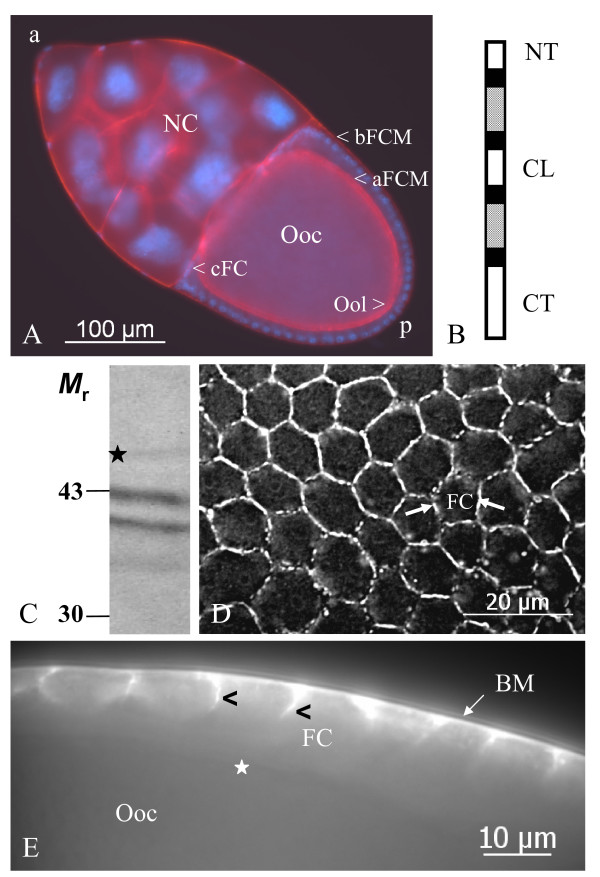
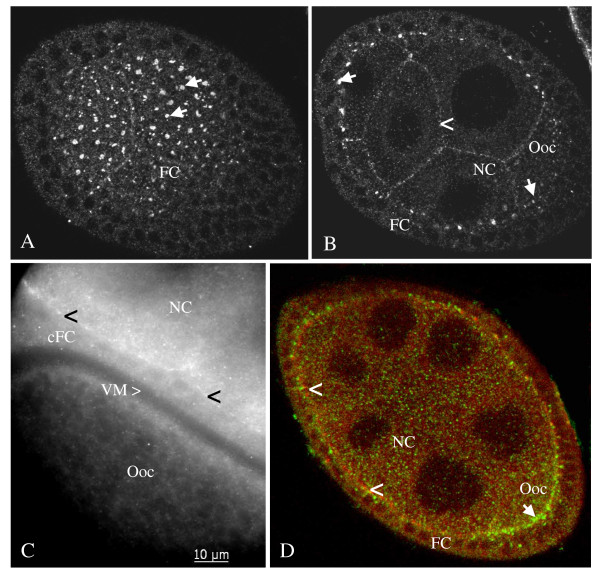
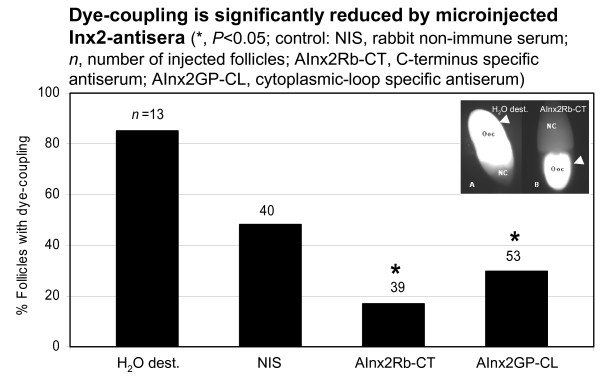
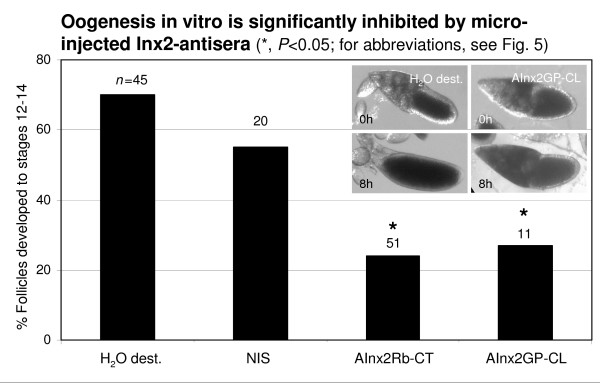
Results: We found innexin 1 predominantly localized to the baso-lateral domain of follicle cells, whereas innexin 2 is positioned apico-laterally as well as apically between follicle cells and germ-line cells. Innexin 3 was observed laterally in follicle cells and also in nurse cells, and innexin 4 was detected in the oolemma up to stage 8 and in nurse-cell membranes up to stage 12. In order to test whether innexins form channels suitable for intercellular communication, we microinjected innexin antibodies in combination with a fluorescent tracer into the oocyte of stage-10 follicles. We found that dye-coupling between oocyte and follicle cells was largely reduced by innexin-2 antibodies directed against the intracellular C-terminus as well as against the intracellular loop. Analyzing in vitro, between stages 10 and 14, the developmental capacities of follicles following microinjections of innexin-2 antibodies revealed defects in follicle-cell differentiation, nurse-cell regression, oocyte growth and choriogenesis.
Conclusion: Our results suggest that all analyzed innexins are involved in the formation of gap junctions in the ovary. While innexins 2 and 3 are colocalized between soma cells, innexins 2 and 4 are colocalized between soma and germ-line cells. Innexin 2 is participating in cell-cell communication via hemichannels residing in the oolemma. It is obvious that gap-junctional communication between germ-line and soma cells is essential for several processes during oogenesis.
Figures


Similar articles
-
Relating proton pumps with gap junctions: colocalization of ductin, the channel-forming subunit c of V-ATPase, with subunit a and with innexins 2 and 3 during Drosophila oogenesis.BMC Dev Biol. 2016 Jul 13;16(1):24. doi: 10.1186/s12861-016-0124-y. BMC Dev Biol. 2016. PMID: 27412523 Free PMC article.
-
Cellular distribution of innexin 1 and 2 gap junctional channel proteins in epithelia of the Drosophila embryo.Cell Commun Adhes. 2003 Jul-Dec;10(4-6):221-5. doi: 10.1080/cac.10.4-6.221.225. Cell Commun Adhes. 2003. PMID: 14681020
-
Gap junction channel protein innexin 2 is essential for epithelial morphogenesis in the Drosophila embryo.Mol Biol Cell. 2004 Jun;15(6):2992-3004. doi: 10.1091/mbc.e04-01-0056. Epub 2004 Mar 26. Mol Biol Cell. 2004. PMID: 15047872 Free PMC article.
-
Innexins: members of an evolutionarily conserved family of gap-junction proteins.Biochim Biophys Acta. 2005 Jun 10;1711(2):225-45. doi: 10.1016/j.bbamem.2004.10.004. Epub 2004 Nov 17. Biochim Biophys Acta. 2005. PMID: 15921654 Review.
-
Intercellular communication: the Drosophila innexin multiprotein family of gap junction proteins.Chem Biol. 2005 May;12(5):515-26. doi: 10.1016/j.chembiol.2005.02.013. Chem Biol. 2005. PMID: 15911372 Review.
Cited by
-
The molecular and immunochemical expression of innexins in the yellow fever mosquito, Aedes aegypti: insights into putative life stage- and tissue-specific functions of gap junctions.Comp Biochem Physiol B Biochem Mol Biol. 2015 May;183:11-21. doi: 10.1016/j.cbpb.2014.11.013. Epub 2015 Jan 10. Comp Biochem Physiol B Biochem Mol Biol. 2015. PMID: 25585357 Free PMC article.
-
Insights into eyestalk ablation mechanism to induce ovarian maturation in the black tiger shrimp.PLoS One. 2011;6(9):e24427. doi: 10.1371/journal.pone.0024427. Epub 2011 Sep 7. PLoS One. 2011. PMID: 21915325 Free PMC article.
-
Ion Channel Contributions to Wing Development in Drosophila melanogaster.G3 (Bethesda). 2019 Apr 9;9(4):999-1008. doi: 10.1534/g3.119.400028. G3 (Bethesda). 2019. PMID: 30733380 Free PMC article.
-
A thermogenetics protocol for detecting gap junction channels in Drosophila egg chambers.STAR Protoc. 2021 Jan 14;2(1):100269. doi: 10.1016/j.xpro.2020.100269. eCollection 2021 Mar 19. STAR Protoc. 2021. PMID: 33490985 Free PMC article.
-
Electrochemical patterns during Drosophila oogenesis: ion-transport mechanisms generate stage-specific gradients of pH and membrane potential in the follicle-cell epithelium.BMC Dev Biol. 2019 Jun 21;19(1):12. doi: 10.1186/s12861-019-0192-x. BMC Dev Biol. 2019. PMID: 31226923 Free PMC article.
References
-
- Spradling A. Developmental genetics of oogenesis. In: Bate M, Martinez Arias A, editor. The Development of Drosophila melanogaster. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press; 1993. pp. 1–70.
-
- Bownes M. Interactions between germ cells and somatic cells in Drosophila melanogaster. Semin Dev Biol. 1994;5:31–42.
-
- Horne-Badovinac S, Bilder D. Mass transit: Epithelial morphogenesis in the Drosophila egg chamber. Dev Dyn. 2005;232:559–574. - PubMed
-
- Schüpbach T, Clifford RJ, Manseau LJ, Price JV. Dorsoventral signaling processes in Drosophila oogenesis. In: Gerhart J, editor. Cell-Cell Interactions in Early Development. New York: Wiley-Liss; 1991. pp. 163–174.
-
- Stevens LM, Nüsslein-Volhard C. Development of the terminal anlagen of the Drosophila embryo depends upon interactions between the germline and the somatic follicle cells. In: Gerhart J, editor. Cell-Cell Interactions in Early Development. New York: Wiley-Liss; 1991. pp. 145–162.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
