

Survival and proliferative roles of erythropoietin beyond the erythroid lineage
- PMID: 19040789
- PMCID: PMC3065109
- DOI: 10.1017/S1462399408000860
Survival and proliferative roles of erythropoietin beyond the erythroid lineage
Abstract
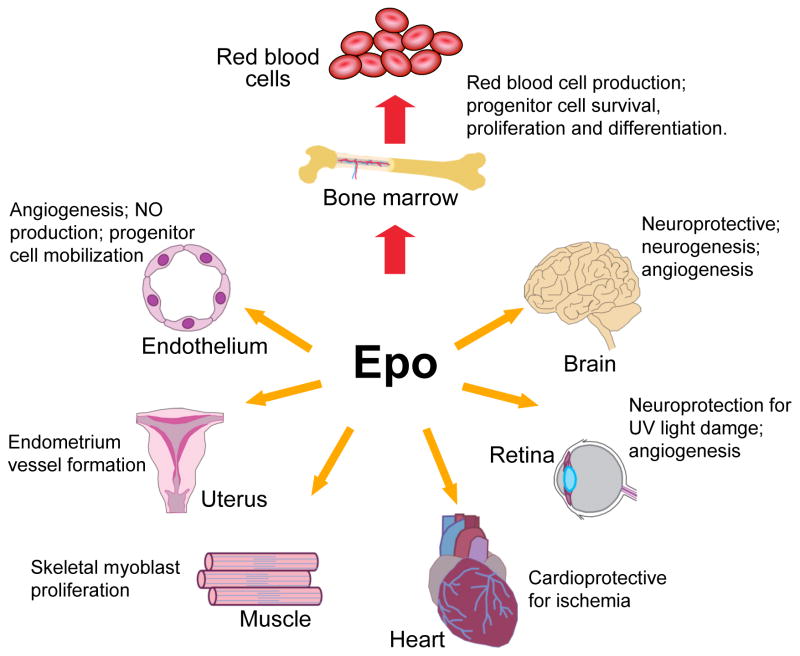
Since the isolation and purification of erythropoietin (EPO) in 1977, the essential role of EPO for mature red blood cell production has been well established. The cloning of the EPO gene and production of recombinant human EPO led to the widespread use of EPO in treating patients with anaemia. However, the biological activity of EPO is not restricted to regulation of erythropoiesis. EPO receptor (EPOR) expression is also found in endothelial, brain, cardiovascular and other tissues, although at levels considerably lower than that of erythroid progenitor cells. This review discusses the survival and proliferative activity of EPO that extends beyond erythroid progenitor cells. Loss of EpoR expression in mouse models provides evidence for the role of endogenous EPO signalling in nonhaematopoietic tissue during development or for tissue maintenance and/or repair. Determining the extent and distribution of receptor expression provides insights into the potential protective activity of EPO in brain, heart and other nonhaematopoietic tissues.
Figures
References
-
- Miyake T, Kung CK, Goldwasser E. Purification of human erythropoietin. J Biol Chem. 1977;252:5558–5564. - PubMed
-
- Jacobs K, et al. Isolation and characterization of genomic and cDNA clones of human erythropoietin. Nature. 1985;313:806–810. - PubMed
-
- Livnah O, et al. Crystallographic evidence for preformed dimers of erythropoietin receptor before ligand activation. Science. 1999;283:987–990. - PubMed
Further reading, resources and contacts
-
- Hodges VM, et al. Pathophysiology of anemia and erythorcytosis. Crit Rev Oncol Hematol. 2007;64:139–158. This review describes anaemia and erythrocytosis in relation to erythropoietin (EPO) levels in diagnosis and treatment. - PubMed
-
- Sasaki R. Pleiotropic functions of erythropoietin. Intern Med. 2003;42:142–149. This review highlights the early observations of EPO production in the central nervous system and reproductive organs, including the author’s pioneering work on EPO expression and function in these tissues. - PubMed
-
- Jelkmann W, et al. The erythropoietin receptor in normal and cancer tissues. Crit Rev Oncol/Hematol. 2008;67:39–61. This paper reviews EPO receptor expression and function in tumour tissues and the implications for administration of EPO to treat anaemia in cancer patients. - PubMed
-
- Sytkowski AJ. Erythropoietin: Blood, Brain and Beyond. Wiley-VCH; Weinheim, Germany: 2004. This book provides an overview of the history, developmental biology, transcription regulation and biochemistry of EPO in the context of erythropoiesis and EPO therapy in anaemia in relation to a variety of clinical conditions. Other topics include EPO in nonhaematopoietic tissues and in elite athletics.
-
-
Websites:
This page from the Information Center for Sickle Cell and Thalassemic Disorders website includes an overview of the relationship between iron and erythropoietin activity in red blood cell production: http://sickle.bwh.harvard.edu/iron_epo.html
-
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
