

Review
doi: 10.1016/j.cell.2008.11.006.
Polarity and differential inheritance--universal attributes of life?
Affiliations
- PMID: 19041746
- PMCID: PMC2844324
- DOI: 10.1016/j.cell.2008.11.006
Item in Clipboard
Review
Polarity and differential inheritance--universal attributes of life?
Cell.
.
Abstract
When and why did cell polarization arise? Recent work in bacteria and yeast suggests that polarization may have evolved to restrict senescence to one daughter during division by enabling the differential segregation of damaged material. In more complex organisms, polarity functions have diversified to permit the differential inheritance of centrosomes, RNAs, proteins, and membranes, which is essential for the generation of diverse cell types from stem cells and for morphogenesis.
Figures
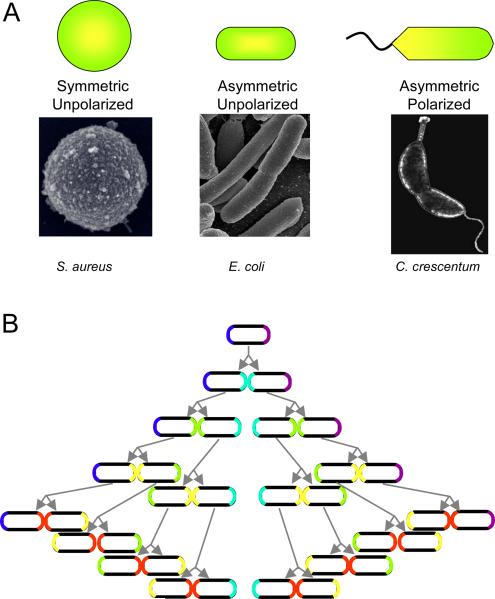
Types of cell polarity. (A) Degrees of cell polarization, with examples. (Electron micrograph of S. aureus by Taeok Bae, Univ. of Chicago) (B) Inheritance of old poles by a symmetrically dividing cell, such as E. coli. The oldest pole is purple. One cell will always retain this pole, while the other pole will always be new. If proteins or other cellular components can recognize old from new poles, they can be segregated asymmetrically during each cell division.
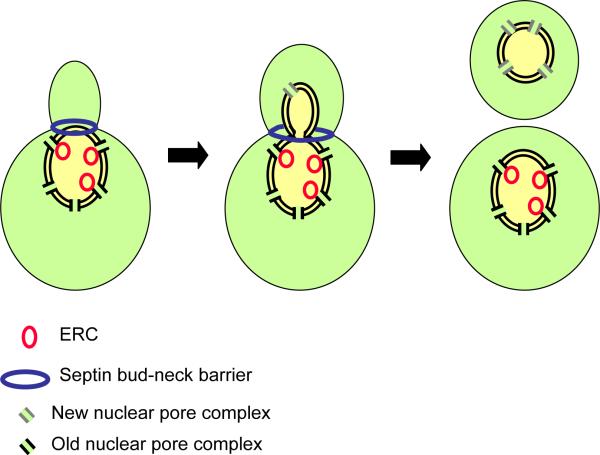
A possible mechanism for the differential inheritance of DNA circles by budding yeast. During mitosis the nucleus penetrates into the bud, but a septin-based diffusion barrier at the bud neck prevents the movement of nuclear pore complexes out of the mother. DNA circles, which contribute to aging, are anchored by the nuclear pore complexes and thereby retained by the mother cell (Shcheprova et al. 2008).
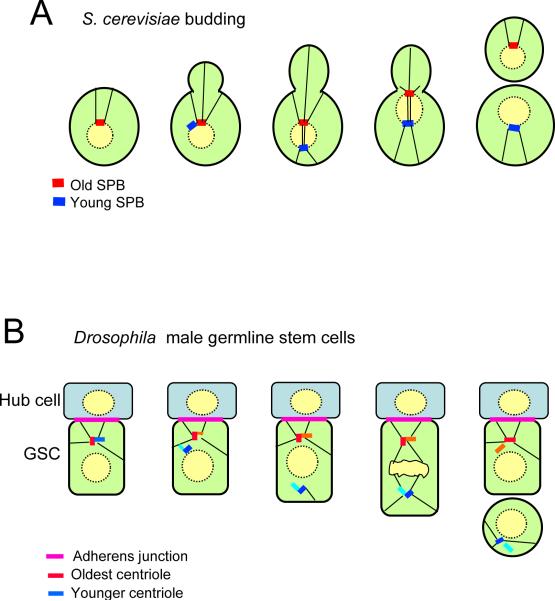
Asymmetric inheritance of spindle pole bodies (SPB) and centrosomes. (A) In budding yeast the old SPB is inherited by the daughter, while the young SPB is retained by the mother. (B) In the germline stem cells of Drosophila, the old centrosome remains in the stem cell, probably by attachment through microtubules to the adherens junction between the stem cell and the hub cell. The differentiating daughter inherits the young centrosome.
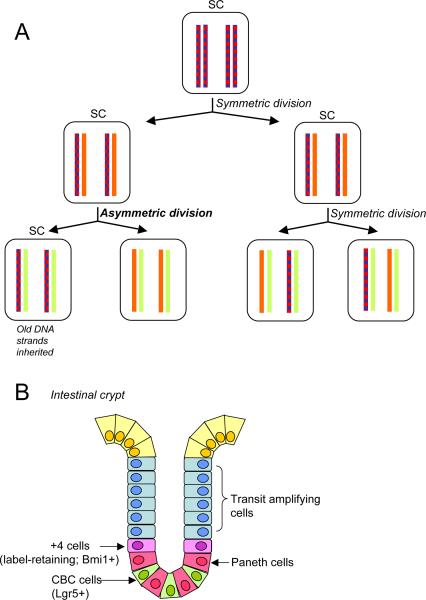
The immortal strand hypothesis. (A) A schematic of differential retention of parental DNA strands by the stem cell. The first division shown is symmetric, leading to amplification of the stem cell population. The second shows one asymmetric division in which the stem cell retains the parental strands; while the other is a classical symmetric division. Blue represents BRdU labeling. (B) Schematic of a small intestinal crypt showing the organization of stem cells and other cell types. In this tissue both the +4 cell that shows BrDU label retention (an attribute of the immortal strand hypothesis) and crypt base columnar cells (CBC), which do not retain label, function as intestinal stem cells.
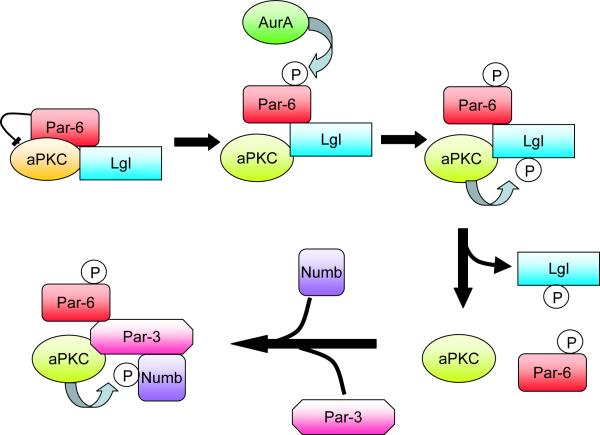
A model for the regulation of Numb phosphorylation by aPKC during Drosophila asymmetric cell divisions. Phosphorylation of the Par-6 polarity protein releases its inhibitory effect on aPKC, which enables the kinase to phosphorylate Lgl. Lgl is then replaced by Par-3 in the complex, which recruits Numb to be phosphorylated. Phosphorylated Numb is released from the cell cortex. Since Par-3 is restricted to the cortex on one side of the cell, Numb becomes restricted to a cortical crescent on the opposite side (Wirtz-Peitz et al., 2008).
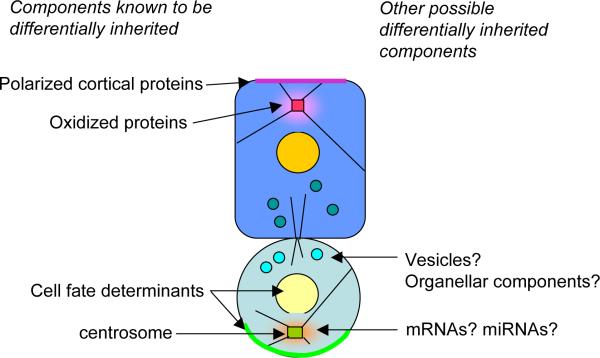
Summary of known and potential cellular components that are differentially inherited during asymmetric stem cell divisions.
References
-
- Ackermann M, Stearns SC, Jenal U. Senescence in a bacterium with asymmetric division. Science. 2003;300:1920. - PubMed
-
- Aguilaniu H, Gustafsson L, Rigoulet M, Nystrom T. Asymmetric inheritance of oxidatively damaged proteins during cytokinesis. Science. 2003;299:1751–1753. - PubMed
-
- Barker MG, Walmsley RM. Replicative ageing in the fission yeast Schizosaccharomyces pombe. Yeast. 1999;15:1511–1518. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
