

Deregulation of scribble promotes mammary tumorigenesis and reveals a role for cell polarity in carcinoma
- PMID: 19041750
- PMCID: PMC3015046
- DOI: 10.1016/j.cell.2008.09.045
Deregulation of scribble promotes mammary tumorigenesis and reveals a role for cell polarity in carcinoma
Abstract
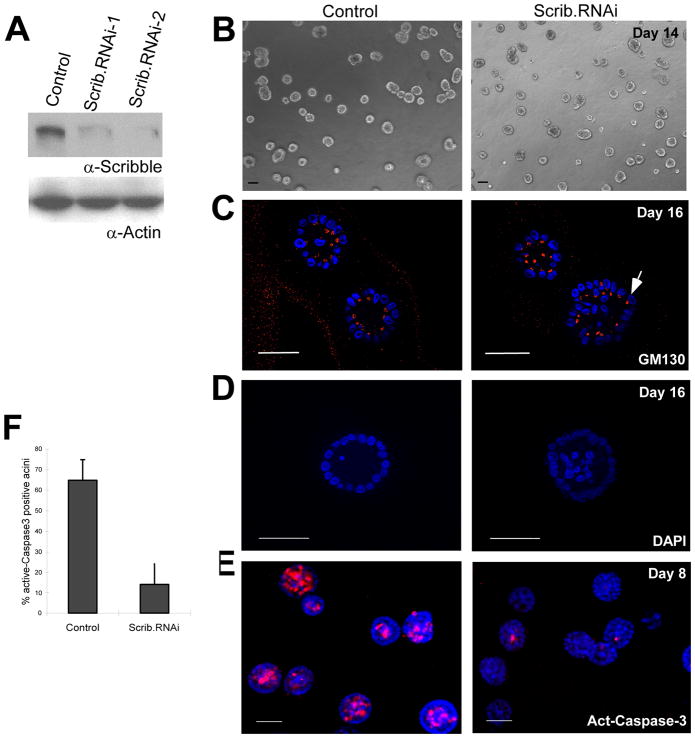
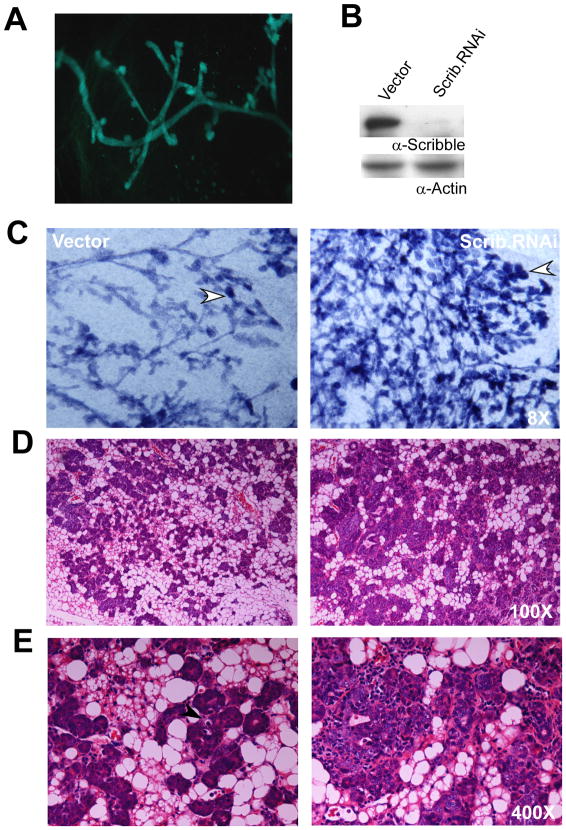
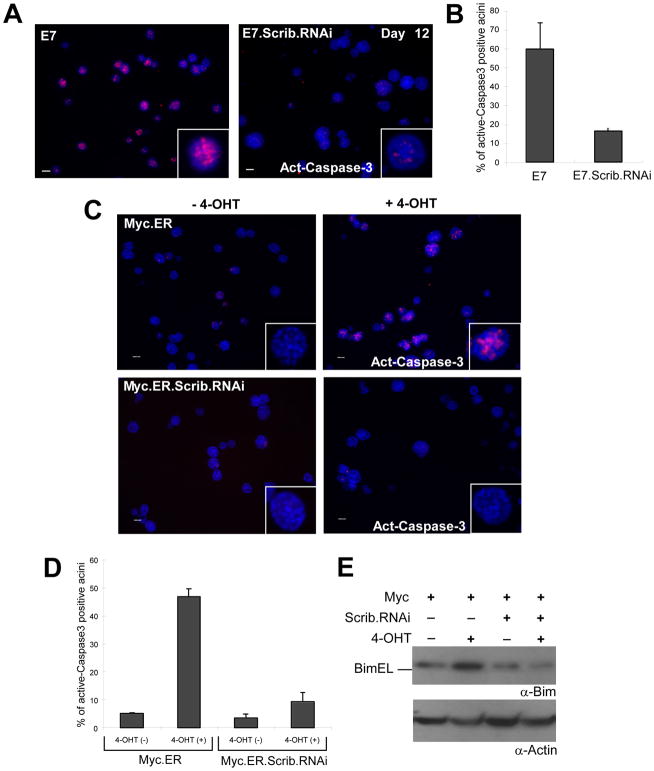
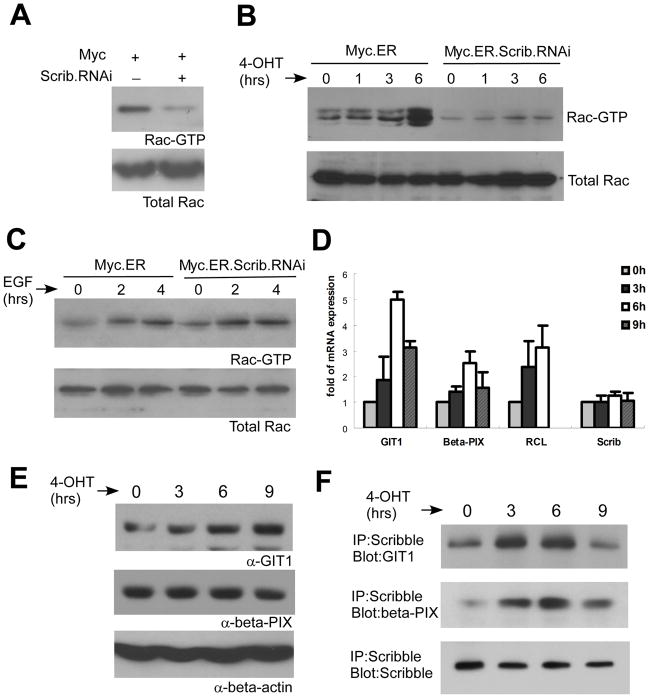
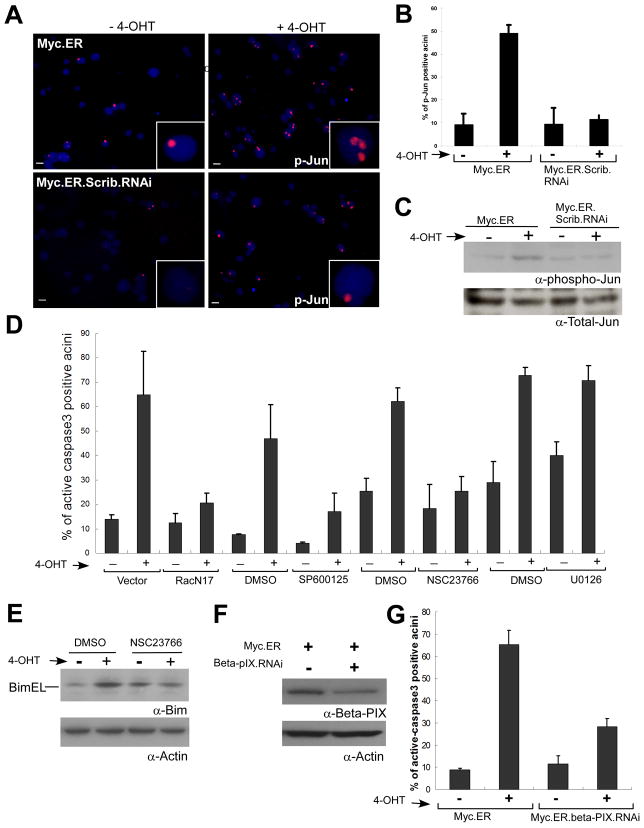
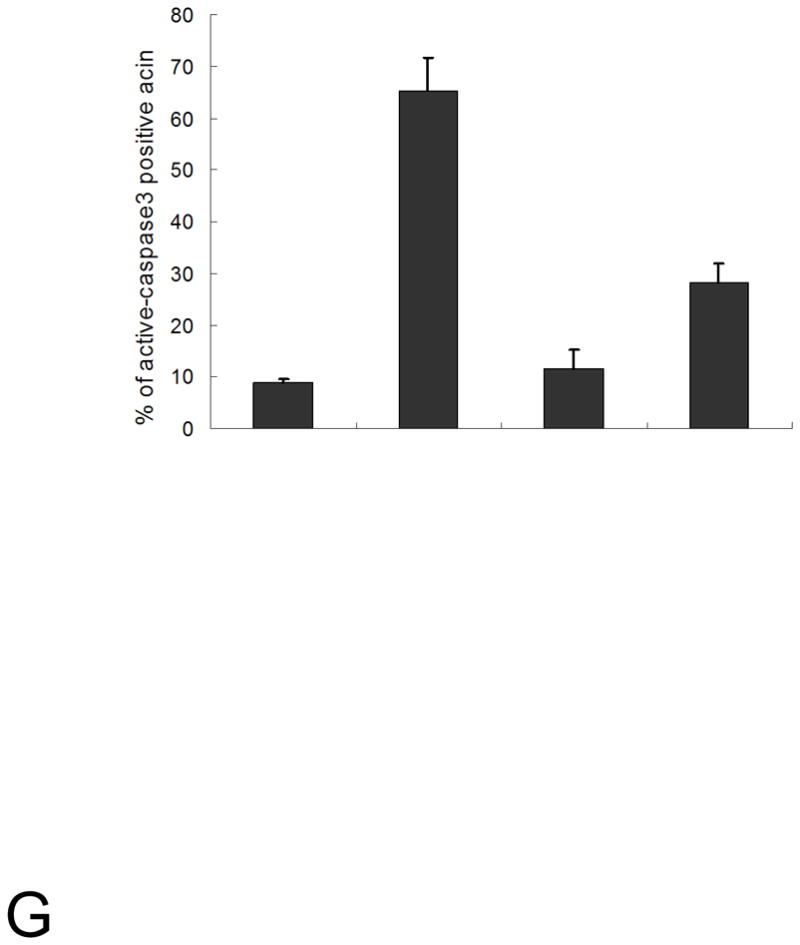
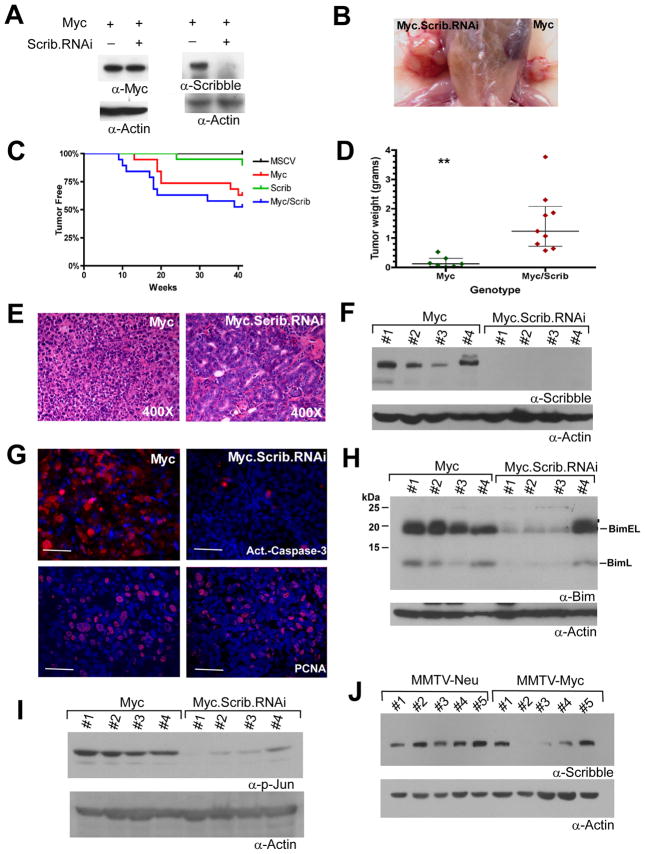
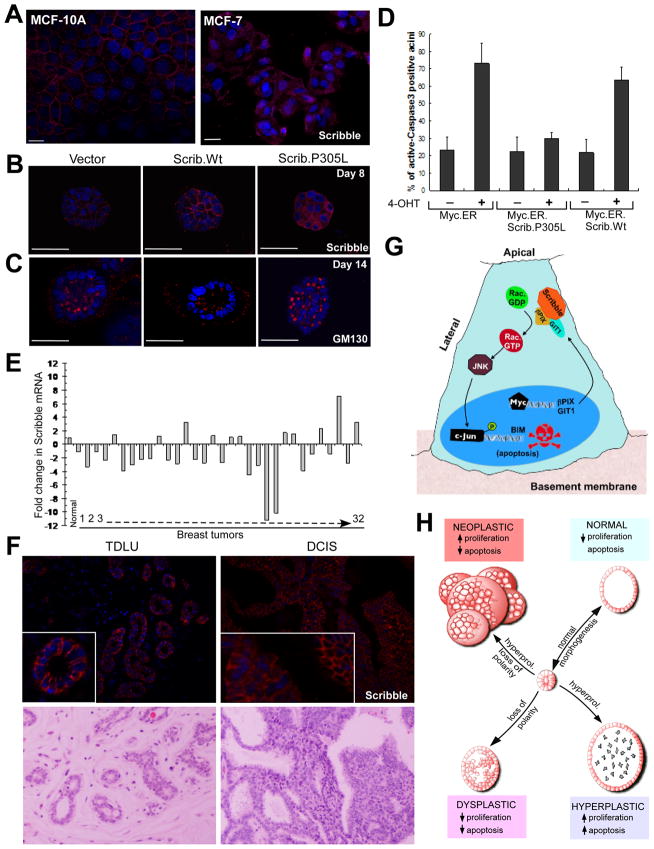
Loss of cell polarity proteins such as Scribble induces neoplasia in Drosophila by promoting uncontrolled proliferation. In mammals, the role that polarity proteins play during tumorigenesis is not well understood. Here, we demonstrate that depletion of Scribble in mammary epithelia disrupts cell polarity, blocks three-dimensional morphogenesis, inhibits apoptosis, and induces dysplasia in vivo that progress to tumors after long latency. Loss of Scribble cooperates with oncogenes such as c-myc to transform epithelial cells and induce tumors in vivo by blocking activation of an apoptosis pathway. Like depletion, mislocalization of Scribble from cell-cell junction was sufficient to promote cell transformation. Interestingly, spontaneous mammary tumors in mice and humans possess both downregulated and mislocalized Scribble. Thus, we demonstrate that scribble inhibits breast cancer formation and that deregulation of polarity pathways promotes dysplastic and neoplastic growth in mammals by disrupting morphogenesis and inhibiting cell death.
Figures
References
-
- Aguilar-Cordova E, Strange R, Young LJ, Billy HT, Gumerlock PH, Cardiff RD. Viral Ha-ras mediated mammary tumor progression. Oncogene. 1991;6:1601–1607. - PubMed
-
- Allred CA, Hilsenback SG, Mohsin SK. Biological features of human premaligant breast disease. In: Harris J, Lippman ME, Morrow M, Osborne K, editors. Diseases of the Breast. Philadelphia: Lippincott Williams & Wilkins; 2004. pp. 507–520.
-
- Allred DC, Mohsin SK, Fuqua SA. Histological and biological evolution of human premalignant breast disease. Endocr Relat Cancer. 2001;8:47–61. - PubMed
-
- Amundadottir LT, Johnson MD, Merlino G, Smith GH, Dickson RB. Synergistic interaction of transforming growth factor alpha and c-myc in mouse mammary and salivary gland tumorigenesis. Cell Growth Differ. 1995;6:737–748. - PubMed
-
- Amundadottir LT, Nass SJ, Berchem GJ, Johnson MD, Dickson RB. Cooperation of TGF alpha and c-Myc in mouse mammary tumorigenesis: coordinated stimulation of growth and suppression of apoptosis. Oncogene. 1996;13:757–765. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
