

Calcitonin gene-related peptide (CGRP) triggers Ca2+ responses in cultured astrocytes and in Bergmann glial cells from cerebellar slices
- PMID: 19046367
- PMCID: PMC4461196
- DOI: 10.1111/j.1460-9568.2008.06514.x
Calcitonin gene-related peptide (CGRP) triggers Ca2+ responses in cultured astrocytes and in Bergmann glial cells from cerebellar slices
Erratum in
- Eur J Neurosci. 2008 Dec;28(12):2550
Abstract
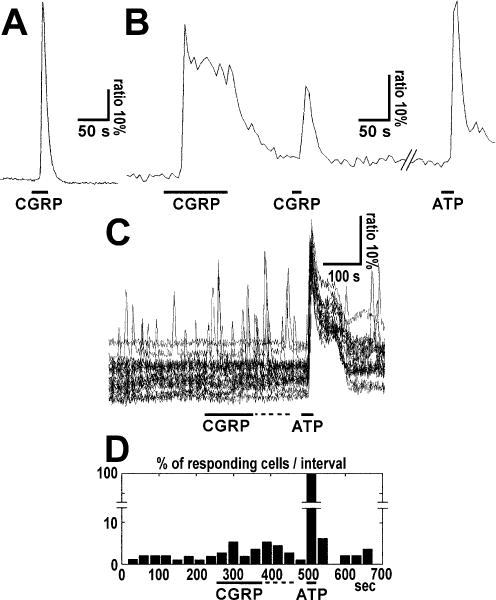
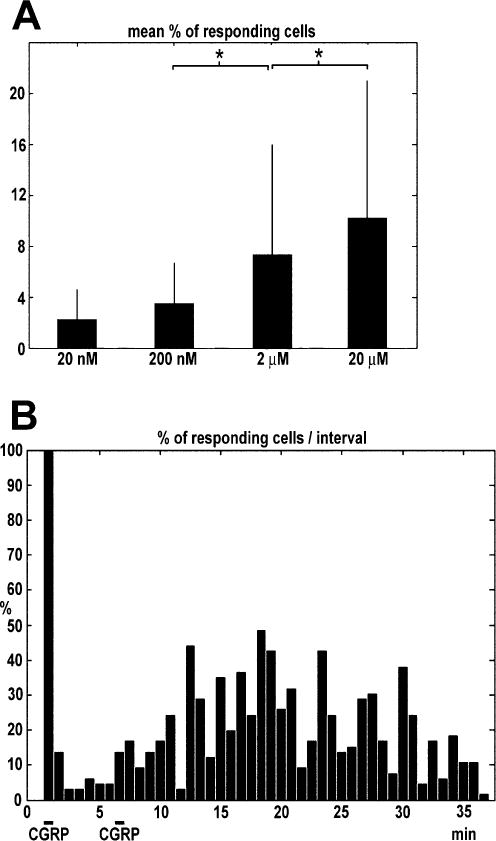
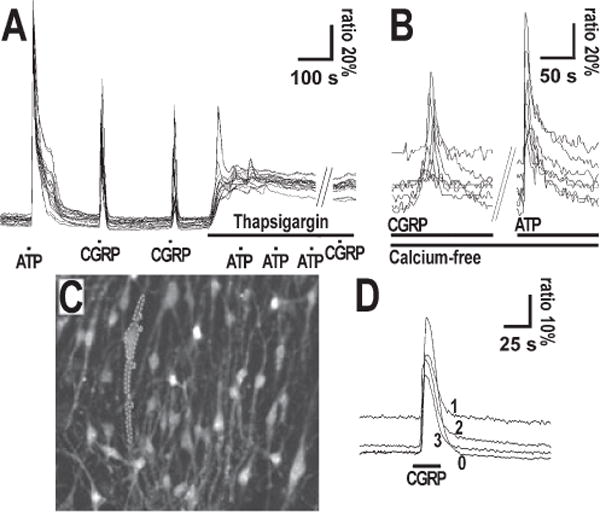
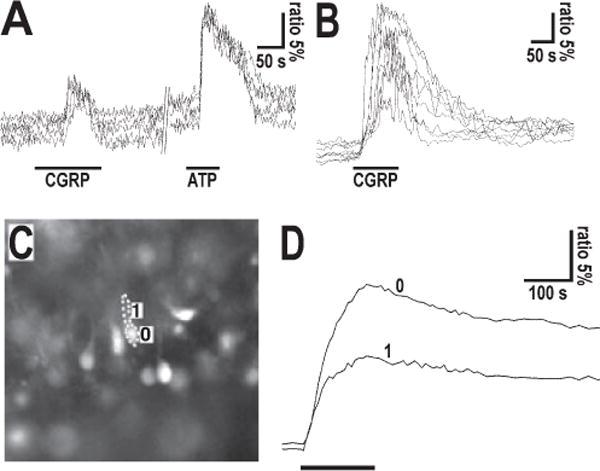
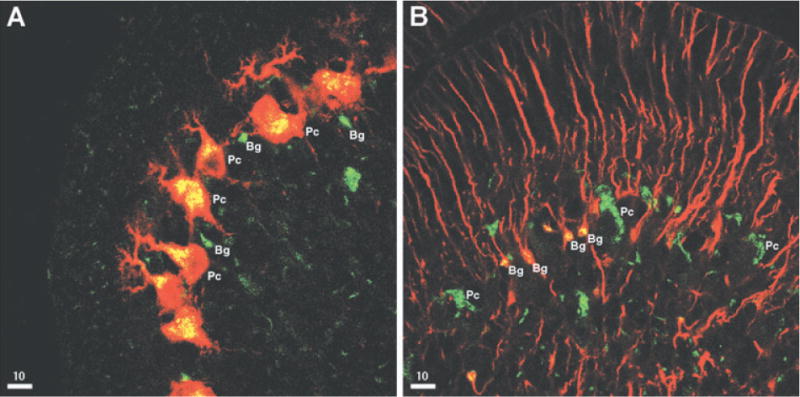
The neuropeptide calcitonin gene-related peptide (CGRP) is transiently expressed in cerebellar climbing fibers during development while its receptor is mainly expressed in astrocytes, in particular Bergmann glial cells. Here, we analyzed the effects of CGRP on astrocytic calcium signaling. Mouse cultured astrocytes from cerebellar or cerebral cortex as well as Bergmann glial cells from acutely isolated cerebellar slices were loaded with the Ca(2+) sensor Fura-2. CGRP triggered transient increases in intracellular Ca(2+) in astrocytes in culture as well as in acute slices. Responses were observed in the concentration range of 1 nm to 1 mm, in both the cell body and its processes. The calcium transients were dependent on release from intracellular stores as they were blocked by thapsigargin but not by the absence of extracellular calcium. In addition, after CGRP application a further delayed transient increase in calcium activity could be observed. Finally, cerebellar astrocytes from neonatal mice expressed receptor component protein, a component of the CGRP receptor, as revealed by immunofluorescence and confocal microscopy. It is thus proposed that the CGRP-containing afferent fibers in the cerebellum (the climbing fibers) modulate calcium in astrocytes by releasing the neuropeptide during development and hence possibly influence the differentiation of Purkinje cells.
Figures
References
-
- Adams JC, Mroz EA, Sewell WF. A possible neurotransmitter role for CGRP in a hair-cell sensory organ. Brain Res. 1987;419:347–351. - PubMed
-
- Aiyar N, Disa J, Stadel JM, Lysko PG. Calcitonin gene-related peptide receptor independently stimulates 3′,5′-cyclic adenosine monophosphate and Ca2+ signaling pathways. Mol Cell Biochem. 1999;197:179–185. - PubMed
-
- Altman J. Morphological development of the rat cerebellum and some of its mechanisms. In: Palay SL, Chan-Palay V, editors. The Cerebellum New Vistas. Springer Verlag; Berlin-Heidelberg: 1982. pp. 8–46.
-
- Armengol JA, Sotelo C. Early dendritic development of Purkinje cells in the rat cerebellum. A light and electron microscopic study using axonal tracing in ‘in vitro’ slices. Dev Brain Res. 1991;64:95–114. - PubMed
-
- Burns DM, Stehno-Bittel L, Kawase T. Calcitonin gene-related peptide elevates calcium and polarizes membrane potential in MG-63 cells by both cAMP-independent and -dependent mechanisms. Am J Physiol Cell Physiol. 2004;287:C457–C467. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
