

E. coli mismatch repair acts downstream of replication fork stalling to stabilize the expanded (GAA.TTC)(n) sequence
- PMID: 19046977
- PMCID: PMC2637364
- DOI: 10.1016/j.mrfmmm.2008.11.003
E. coli mismatch repair acts downstream of replication fork stalling to stabilize the expanded (GAA.TTC)(n) sequence
Abstract
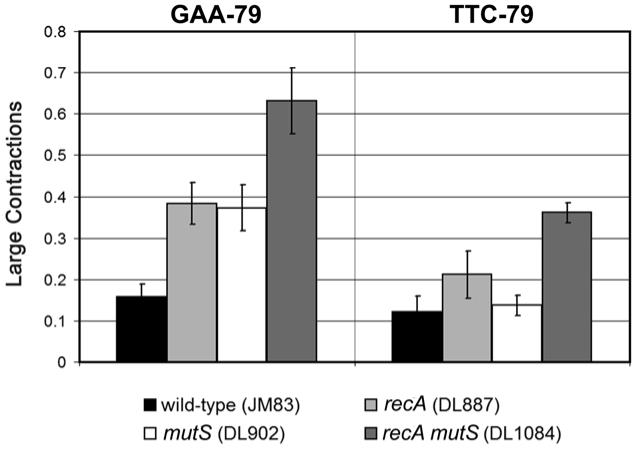
Expanded triplet repeat sequences are known to cause at least 16 inherited neuromuscular diseases. In addition to short-length changes, expanded triplet repeat tracts frequently undergo large changes, often amounting to hundreds of base-pairs. Such changes might occur when template or primer slipping creates insertion/deletion loops (IDLs), which are normally repaired by the mismatch repair system (MMR). However, in prokaryotes and eukaryotes, MMR promotes large changes in the length of (CTG.CAG)(n) sequences, the motif most commonly associated with human disease. We tested the effect of MMR on instability of the expanded (GAA.TTC)(n) sequence, which causes Friedreich ataxia, by comparing repeat instability in wild-type and MMR-deficient strains of Escherichia coli. As expected, the prevalence of small mutations increased in the MMR-deficient strains. However, the prevalence of large contractions increased in the MMR mutants specifically when GAA was the lagging strand template, the orientation in which replication fork stalling is known to occur. After hydroxyurea-induced stalling, both orientations of replication showed significantly more large contractions in MMR mutants than in the wild-type, suggesting that fork stalling may be responsible for the large contractions. Deficiency of MMR promoted large contractions independently of RecA status, a known determinant of (GAA.TTC)(n) instability. These data suggest that two independent mechanisms act in response to replication stalling to prevent instability of the (GAA.TTC)(n) sequence in E. coli, when GAA serves as the lagging strand template: one that is dependent on RecA-mediated restart of stalled forks, and another that is dependent on MMR-mediated repair of IDLs. While MMR destabilizes the (CTG.CAG)(n) sequence, it is involved in stabilization of the (GAA.TTC)(n) sequence. The role of MMR in triplet repeat instability therefore depends on the repeat sequence and the orientation of replication.
Figures



References
-
- Jiricny J. The multifaceted mismatch-repair system. Nat. Rev. Mol. Cell Biol. 2006;7:335–346. - PubMed
-
- Dohet C, Dzidic S, Wagner R, Radman M. Large non-homology in heteroduplex DNA is processed differently than single base pair mismatches. Mol. Gen. Genet. 1987;206:181–184. - PubMed
-
- Fang WH, Wang BJ, Wang CH, Lee SJ, Chang YT, Chuang YK, Lee CN. DNA loop repair by Escherichia coli cell extracts. J. Biol. Chem. 2003;278:22446–22452. - PubMed
-
- Peltomaki P. Role of DNA mismatch repair defects in the pathogenesis of human cancer. J. Clin. Oncol. 2003;21:1174–1179. - PubMed
-
- Lahue RS, Au KG, Modrich P. DNA mismatch correction in a defined system. Science. 1989;245:160–164. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
