

A zinc-dependent adhesion module is responsible for intercellular adhesion in staphylococcal biofilms
- PMID: 19047636
- PMCID: PMC2592360
- DOI: 10.1073/pnas.0807717105
A zinc-dependent adhesion module is responsible for intercellular adhesion in staphylococcal biofilms
Abstract
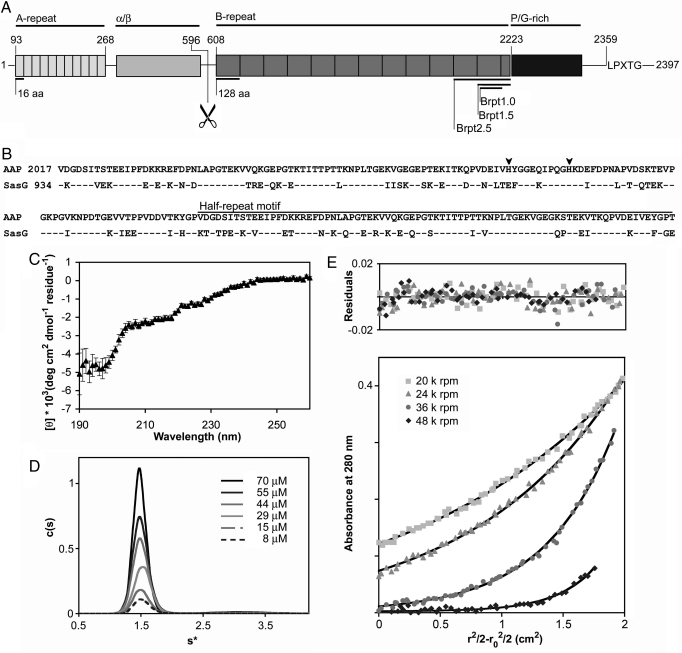
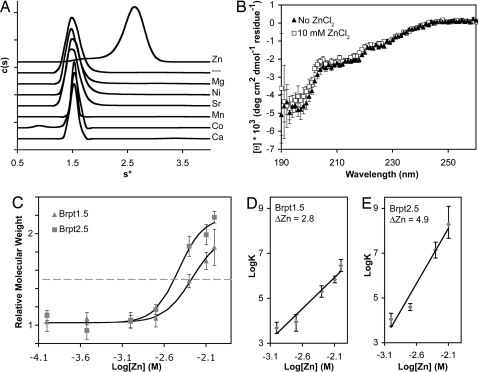
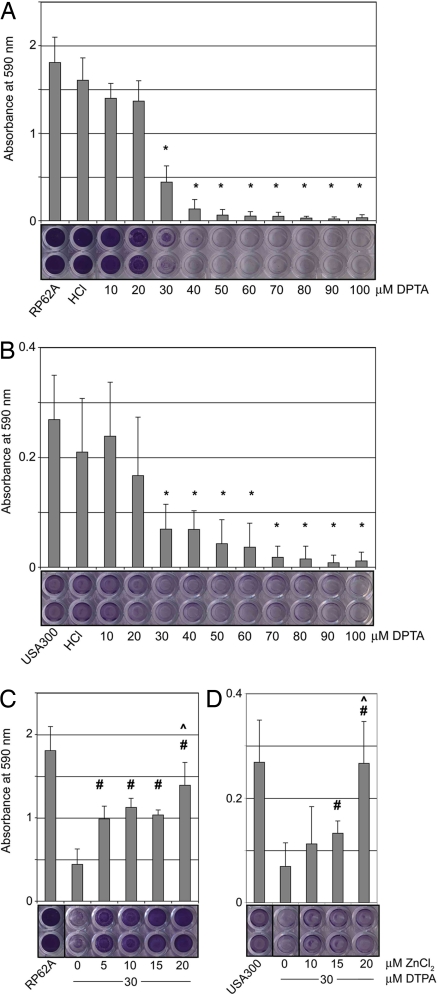
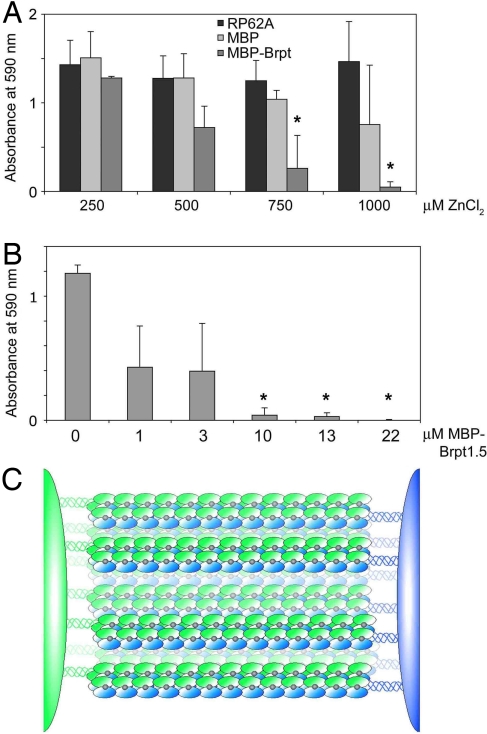
Hospital-acquired bacterial infections are an increasingly important cause of morbidity and mortality worldwide. Staphylococcal species are responsible for the majority of hospital-acquired infections, which are often complicated by the ability of staphylococci to grow as biofilms. Biofilm formation by Staphylococcus epidermidis and Staphylococcus aureus requires cell-surface proteins (Aap and SasG) containing sequence repeats known as G5 domains; however, the precise role of these proteins in biofilm formation is unclear. We show here, using analytical ultracentrifugation (AUC) and circular dichroism (CD), that G5 domains from Aap are zinc (Zn(2+))-dependent adhesion modules analogous to mammalian cadherin domains. The G5 domain dimerizes in the presence of Zn(2+), incorporating 2-3 Zn(2+) ions in the dimer interface. Tandem G5 domains associate in a modular fashion, suggesting a "zinc zipper" mechanism for G5 domain-based intercellular adhesion in staphylococcal biofilms. We demonstrate, using a biofilm plate assay, that Zn(2+) chelation specifically prevents biofilm formation by S. epidermidis and methicillin-resistant S. aureus (MRSA). Furthermore, individual soluble G5 domains inhibit biofilm formation in a dose-dependent manner. Thus, the complex three-dimensional architecture of staphylococcal biofilms results from the self-association of a single type of protein domain. Surface proteins with tandem G5 domains are also found in other bacterial species, suggesting that this mechanism for intercellular adhesion in biofilms may be conserved among staphylococci and other Gram-positive bacteria. Zn(2+) chelation represents a potential therapeutic approach for combating biofilm growth in a wide range of bacterial biofilm-related infections.
Conflict of interest statement
Conflict of interest statement: D.G.C., C.C.B., and A.B.H. have filed a provisional patent application based on the work in this paper.
Figures
References
-
- McCaughey B. Unnecessary deaths: The human and financial costs of hospital infections. Committee to Reduce Infection Deaths. 2006:1–67.
-
- National Nosocomial Infections Surveillance (NNIS) System Report. National Center for Infectious Diseases, Center for Disease Control and Prevention. 1998:1–26.
-
- Costerton JW, Stewart PS, Greenberg EP. Bacterial biofilms: A common cause of persistent infections. Science. 1999;284:1318–1322. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
