

Transglutaminase-1 regulates renal epithelial cell proliferation through activation of Stat-3
- PMID: 19049964
- PMCID: PMC2631946
- DOI: 10.1074/jbc.M808396200
Transglutaminase-1 regulates renal epithelial cell proliferation through activation of Stat-3
Abstract
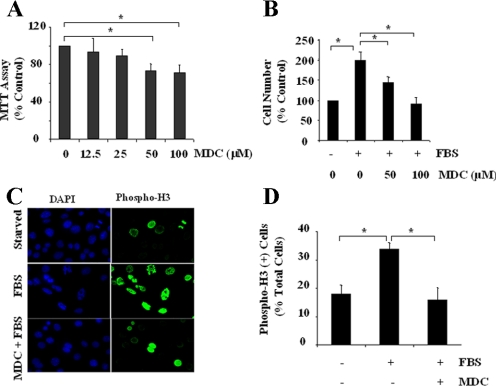
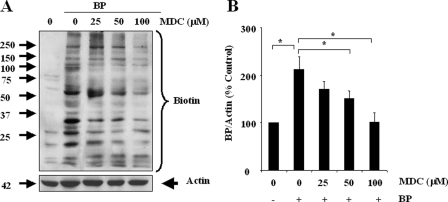
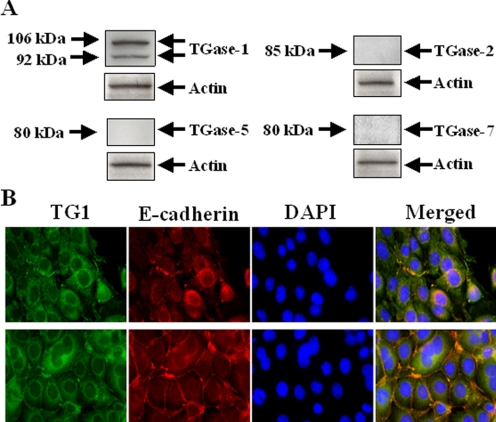
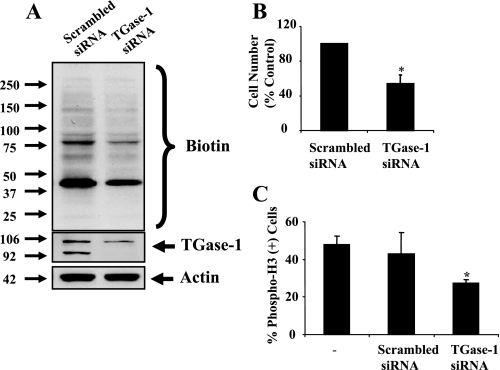
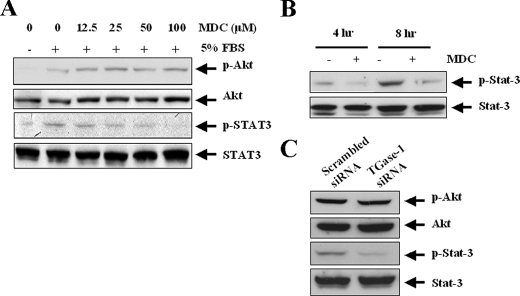
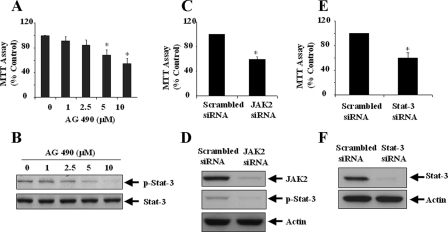
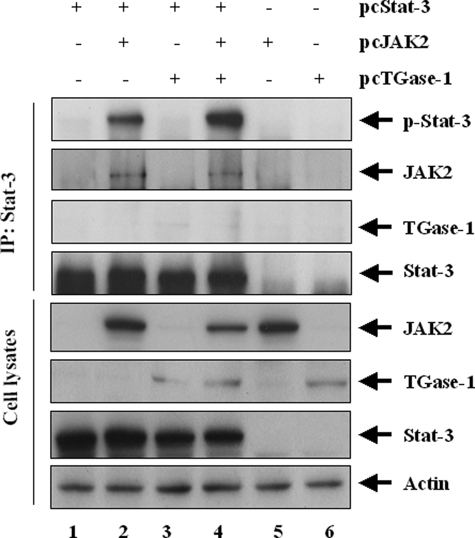
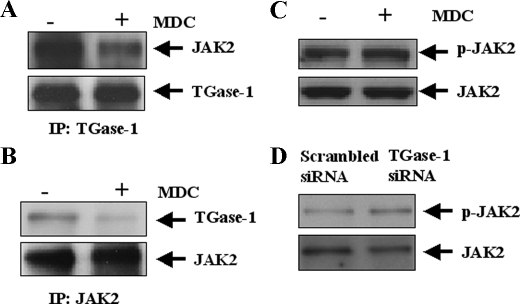
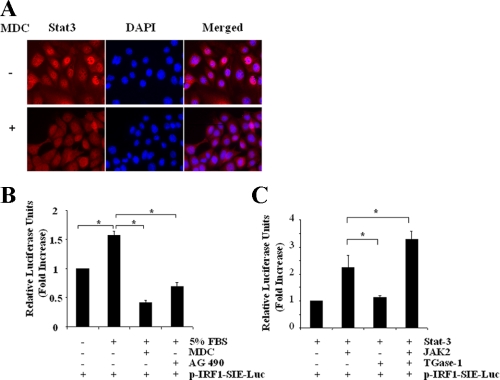
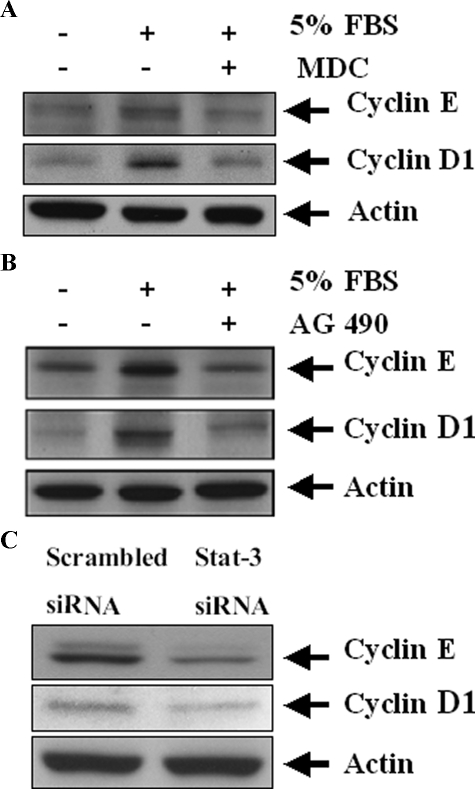
Transglutaminase-1 (TGase-1) is a Ca(2+)-dependent enzyme capable of cross-linking a variety of proteins and promoting wound healing in the skin. In this study, we examined the role of TGase-1 in proliferation of renal proximal tubular cells (RPTC). TGase-1, but not TGase-2, -5, and -7, was expressed in RPTC. Treatment with monodansylcadarevine (MDC), a selective TGase inhibitor or down-regulation of TGase-1 with small interfering RNA (siRNA) decreased RPTC proliferation. Proliferation of RPTC was accompanied by activation of Akt and Stat-3 (signal transducer and activator of transcription-3). Treatment with MDC or TGase-1 siRNA decreased Stat-3 but not Akt phosphorylation. Further studies showed that the Janus-activated kinase 2 (JAK2) mediates phosphorylation of Stat-3, and knockdown of either JAK2 or Stat-3 by siRNA decreased RPTC proliferation. However, inhibition of TGase-1 decreased phosphorylation of Stat-3 but not JAK2. Overexpression of Stat-3, JAK2, and/or TGase-1 in RPTC revealed that JAK2 is indispensable for TGase-1 to induce Stat-3 phosphorylation and TGase-1 potentiates JAK2-induced Stat-3 phosphorylation. Consistent with these observations, we found that inhibition of TGase-1 and the JAK2-Stat-3 signaling pathway decreased the transcriptional activity of Stat-3 and expression of the Stat-3-targeted genes, cyclin D1 and cyclin E. Conversely, overexpresssion of TGase-1 enhanced the JAK2-dependent transcriptional activity of Stat-3. Finally, TGase-1 was found to interact with JAK2, and this interaction was inhibited by MDC. These results demonstrate that TGase-1 plays an important role in regulation of renal epithelial cell proliferation through the JAK2-Stat-3 signaling pathway.
Figures
References
-
- Lorand, L., and Graham, R. M. (2003) Nat. Rev. Mol. Cell. Biol. 4, 140-156 - PubMed
-
- Hiiragi, T., Sasaki, H., Nagafuchi, A., Sabe, H., Shen, S. C., Matsuki, M., Yamanishi, K., and Tsukita, S. (1999) J. Biol. Chem. 274, 34148-34154 - PubMed
-
- Martinet, N., Bonnard, L., Regnault, V., Picard, E., Burke, L., Siat, J., Grosdidier, G., Martinet, Y., and Vignaud, J. M. (2003) Am. J. Respir. Cell Mol. Biol. 28, 428-435 - PubMed
-
- Fesus, L., and Piacentini, M. (2002) Trends Biochem. Sci. 27, 534-539 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
