

doi: 10.1128/JVI.01888-08.
Epub 2008 Dec 3.
Tight junction proteins claudin-1 and occludin control hepatitis C virus entry and are downregulated during infection to prevent superinfection
Affiliations
- PMID: 19052094
- PMCID: PMC2643775
- DOI: 10.1128/JVI.01888-08
Item in Clipboard
Tight junction proteins claudin-1 and occludin control hepatitis C virus entry and are downregulated during infection to prevent superinfection
J Virol.
2009 Feb.
Abstract
A tight junction (TJ) protein, claudin-1 (CLDN1), was identified recently as a key factor for hepatitis C virus (HCV) entry. Here, we show that another TJ protein, occludin, is also required for HCV entry. Mutational study of CLDN1 revealed that its tight junctional distribution plays an important role in mediating viral entry. Together, these data support the model in which HCV enters liver cells from the TJ. Interestingly, HCV infection of Huh-7 hepatoma cells downregulated the expression of CLDN1 and occludin, preventing superinfection. The altered TJ protein expression may contribute to the morphological and functional changes observed in HCV-infected hepatocytes.
Figures
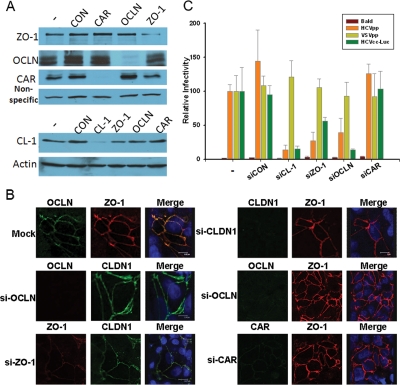
Depletion of OCLN blocks HCV entry. Huh7.5.1 cells (2 × 105) were transfected with the indicated siRNA oligonucleotides (made by Dharmacon, Ambion, and IDTDNA) at the final concentration of 40 nM by the RNAiMax reagent (Invitrogen) for 24 h. Cells were incubated for an additional 24 h, and the specific knockdown of each protein was verified by immunoblotting (A) and confocal microscopy (B). (C) Cells derived from the steps in panel A were infected with the indicated viruses for 2 h and further incubated for 24 to 48 h prior to the luciferase assay. Data plotted are the means ± standard deviations of the results. Bald virus was packaged using the same HIV core construct without the Env-encoding gene, and HCVcc-Luc represents an HCVcc virus with a luciferase gene inserted into its genome. The results shown are representative of three independent experiments.

OCLN coprecipitates with HCV E2 from HCVcc-infected Huh7.5.1 cells. Huh-7.5.1 cells (2 × 109) were infected by JFH1-AM2 (sample 1) and JFH1-FlagE2-AM2 (sample 2) viruses. On day 3 postinfection, cells were trypsinized and washed with ice-cold phosphate-buffered saline and then Dounce homogenized in 10 ml of immunoprecipitation (IP) buffer (20 mM HEPES [pH 7.5], 150 mM NaCl, 1 mM dithiothreitol, 1 mM EDTA, 0.5% NP-40, 5 mM β-glycerophosphate) supplemented with a protease inhibitor cocktail (Sigma). Centrifugation-cleared lysates were then subjected to IP with 50 μl anti-Flag M2 affinity resin (Sigma). After four washes with the IP buffer, bound proteins were eluted with Flag peptide (100 μg/ml; Sigma) in 100 μl Tris-buffered saline. A total of 0.1% of input samples and 7.5 μl of pull-down products were loaded for Western blot analysis. Flag-E2, OCLN, and CAR were detected by their corresponding antibodies.

Junctional CLDN1 is important for HCV entry. (A) 293T cells, free of endogenous CLDN1 (19), were transfected with the indicated CLDN1 constructs (all fused with green fluorescent protein [GFP]). The subcellular distribution of each fusion protein (green) was examined by confocal microscopy. Nuclei were stained with Draq5 (blue). Images shown are XY, XZ, and YZ planes derived from the three-dimensional scanning. (B) A total of 1 × 106 293T cells were transfected with 1 μg of the constructs that were used in panel A. Twenty-four hours after transfection, cells were stained with an anti-CLDN1 antibody (MAB4618; R&D Systems) which specifically recognizes surface CLDN1. Total CLDN1 expression was first determined by measuring the GFP expression on a flow cytometer. Gated GFP-positive cells (blue) were then quantified for surface CLDN1 expression (magenta). The percentage of positive cells and the mean fluorescence intensity are indicated in the figure. (C) 293T cells were transfected with the indicated constructs and further spin-infected by HCVpp for the luciferase assay. Normalized luciferase activity was plotted. Data shown are the means ± standard deviations of the results. The results shown are representative of four independent experiments.

HCV infection downregulates CLDN1/OCLN expression. (A) Naïve Huh7 cells were infected with JFH1 HCVcc (MOI, 0.1) for 5 days or left uninfected. Total cellular CLDN1 and OCLN were determined by Western blotting. (B) The live cells from panel A were analyzed for surface CLDN1 expression by flow cytometry. The rat anti-CLDN1 antibody (MAB4618; R&D Systems) that specifically recognizes surface CLDN1 was used. (C) The cells from panel A were infected with HCVpp to determine the susceptibility to superinfection. (D) 293T cells seeded in a 12-well plate were transfected with combinations of 0.5 μg of CLDN1- or OCLN-expressing plasmid with 0.5 μg of plasmids expressing the indicated viral proteins. Forty-eight hours after transfection, the levels of all proteins were quantified by Western blotting. β-Actin was determined for loading control. FL Core represents full-length HCV Core protein. The truncated Core protein does not have the first 50 amino acids. The position of each individual viral protein is indicated with a star sign. Except for E1 and E2, all proteins were N-terminally Flag-tagged for simple detection by anti-Flag antibody. E1 and E2 were detected by A4 and H52 antibodies, respectively. E1 and E2 are derived from genotype 1a (H77), and other viral proteins were derived from HCV genotype 2a (JFH1).
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
