

Carbonic anhydrase II-positive pancreatic cells are progenitors for both endocrine and exocrine pancreas after birth
- PMID: 19052237
- PMCID: PMC2604974
- DOI: 10.1073/pnas.0805803105
Carbonic anhydrase II-positive pancreatic cells are progenitors for both endocrine and exocrine pancreas after birth
Abstract
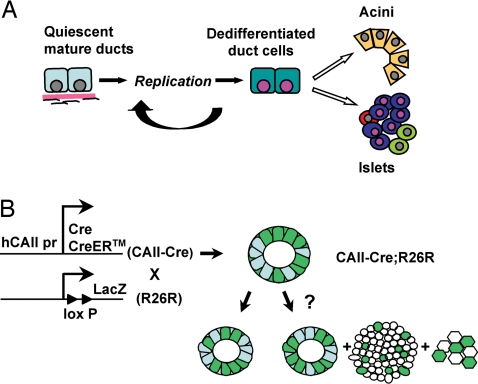
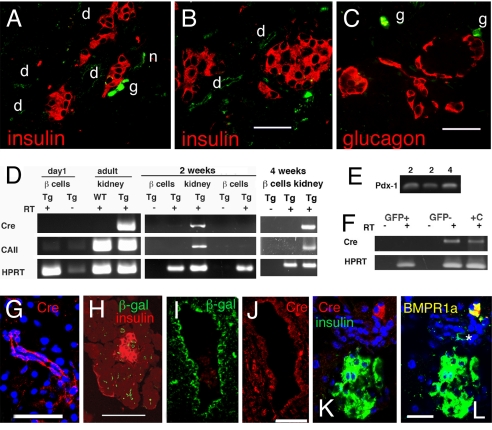
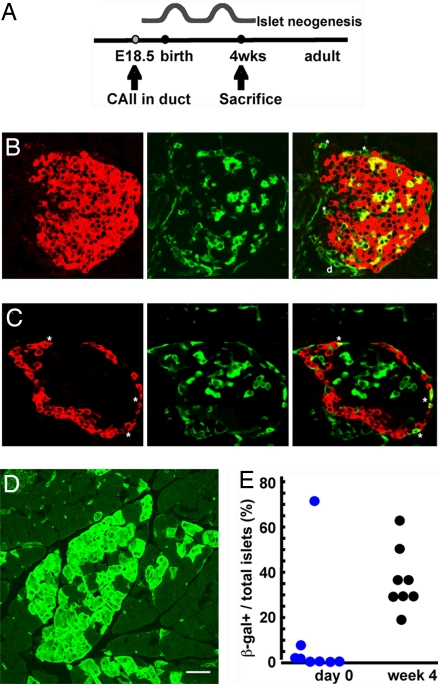
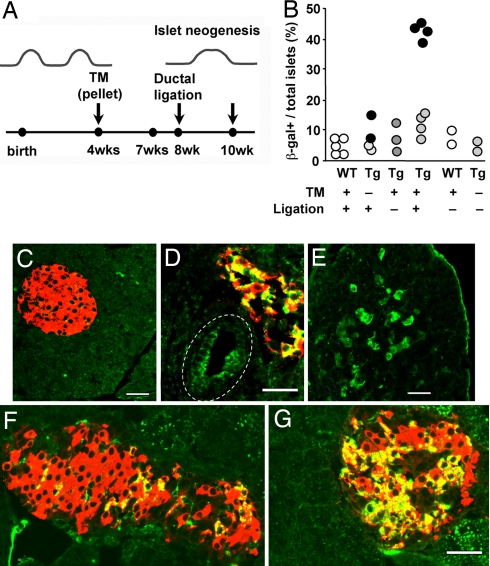
The regenerative process in the pancreas is of particular interest because diabetes results from an inadequate number of insulin-producing beta cells and pancreatic cancer may arise from the uncontrolled growth of progenitor/stem cells. Continued and substantial growth of islet tissue occurs after birth in rodents and humans, with additional compensatory growth in response to increased demand. In rodents there is clear evidence of pancreatic regeneration after some types of injury, with proliferation of preexisting differentiated cell types accounting for some replacement. Additionally, neogenesis or the budding of new islet cells from pancreatic ducts has been reported, but the existence and identity of a progenitor cell have been debated. We hypothesized that the progenitor cells are duct epithelial cells that after replication undergo a regression to a less differentiated state and then can form new endocrine and exocrine pancreas. To directly test whether ductal cells serve as pancreatic progenitors after birth and give rise to new islets, we generated transgenic mice expressing human carbonic anhydrase II (CAII) promoter: Cre recombinase (Cre) or inducible CreER(TM) to cross with ROSA26 loxP-Stop-loxP LacZ reporter mice. We show that CAII-expressing cells within the pancreas act as progenitors that give rise to both new islets and acini normally after birth and after injury (ductal ligation). This identification of a differentiated pancreatic cell type as an in vivo progenitor of all differentiated pancreatic cell types has implications for a potential expandable source for new islets for replenishment therapy for diabetes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Butler AE, et al. Beta-cell deficit and increased beta-cell apoptosis in humans with type 2 diabetes. Diabetes. 2003;52:102–110. - PubMed
-
- Li C, et al. Identification of pancreatic cancer stem cells. Cancer Res. 2007;67:1030–1037. - PubMed
-
- Bonner-Weir S, et al. The pancreatic ductal epithelium serves as a potential pool of progenitor cells. Pediatr Diabetes. 2005;5:15–22. - PubMed
-
- Wang RN, Kloppel G, Bouwens L. Duct- to islet-cell differentiation and islet growth in the pancreas of duct-ligated adult rats. Diabetologia. 1995;38:1405–1411. - PubMed
-
- Brockenbrough JS, Weir GC, Bonner-Weir S. Discordance of exocrine and endocrine growth after 90% pancreatectomy in rats. Diabetes. 1988;37:232–236. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
