

PcG proteins, DNA methylation, and gene repression by chromatin looping
- PMID: 19053175
- PMCID: PMC2592355
- DOI: 10.1371/journal.pbio.0060306
PcG proteins, DNA methylation, and gene repression by chromatin looping
Abstract
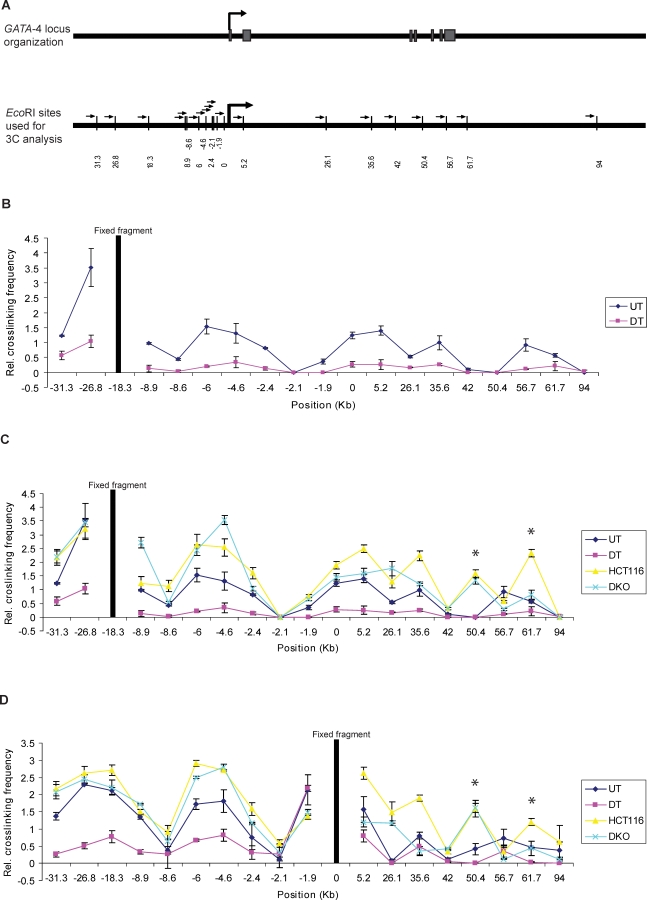
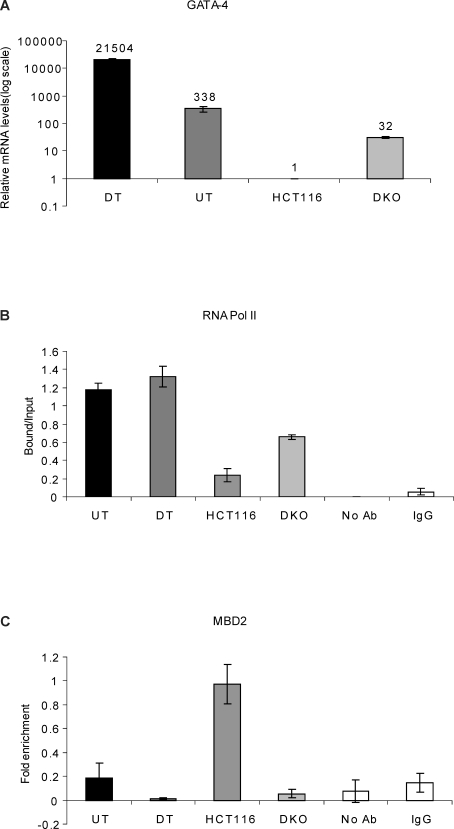
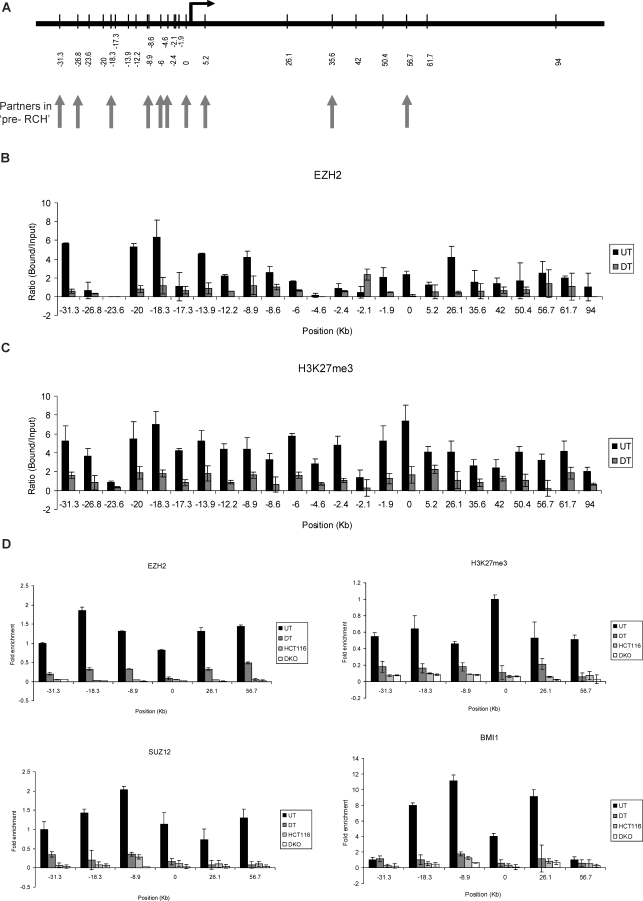
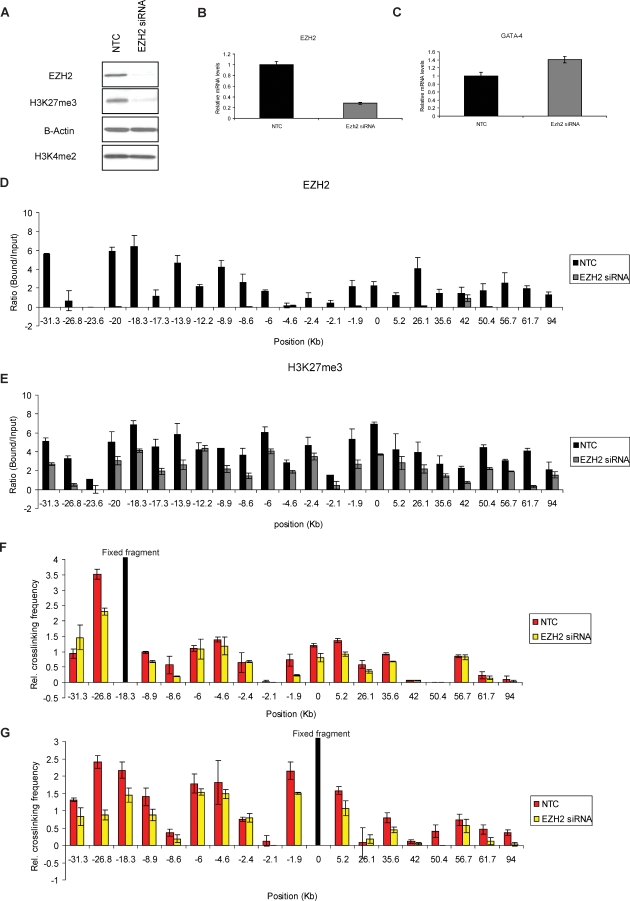
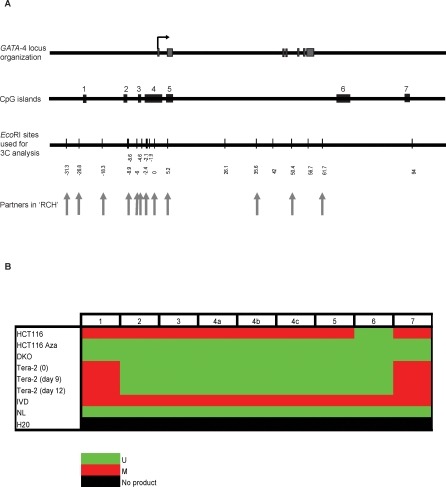
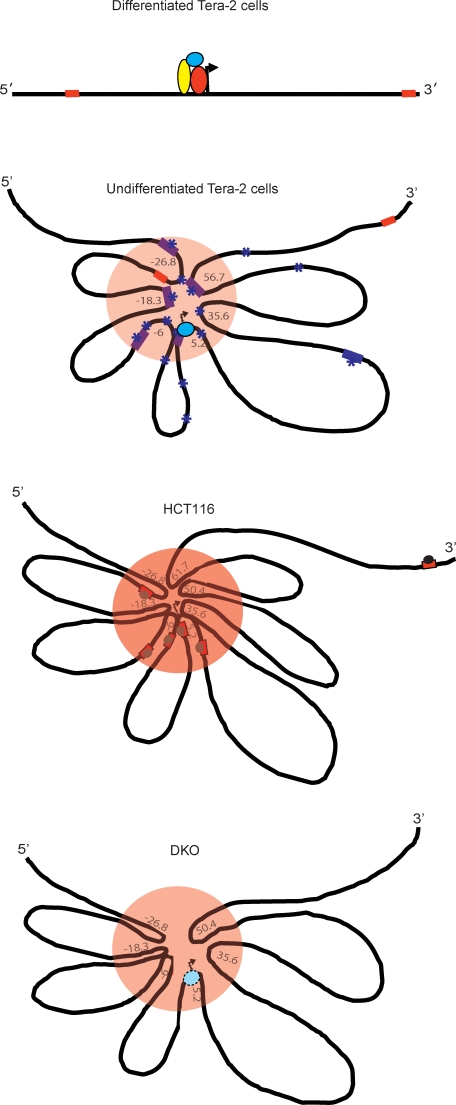
Many DNA hypermethylated and epigenetically silenced genes in adult cancers are Polycomb group (PcG) marked in embryonic stem (ES) cells. We show that a large region upstream ( approximately 30 kb) of and extending approximately 60 kb around one such gene, GATA-4, is organized-in Tera-2 undifferentiated embryonic carcinoma (EC) cells-in a topologically complex multi-loop conformation that is formed by multiple internal long-range contact regions near areas enriched for EZH2, other PcG proteins, and the signature PcG histone mark, H3K27me3. Small interfering RNA (siRNA)-mediated depletion of EZH2 in undifferentiated Tera-2 cells leads to a significant reduction in the frequency of long-range associations at the GATA-4 locus, seemingly dependent on affecting the H3K27me3 enrichments around those chromatin regions, accompanied by a modest increase in GATA-4 transcription. The chromatin loops completely dissolve, accompanied by loss of PcG proteins and H3K27me3 marks, when Tera-2 cells receive differentiation signals which induce a approximately 60-fold increase in GATA-4 expression. In colon cancer cells, however, the frequency of the long-range interactions are increased in a setting where GATA-4 has no basal transcription and the loops encompass multiple, abnormally DNA hypermethylated CpG islands, and the methyl-cytosine binding protein MBD2 is localized to these CpG islands, including ones near the gene promoter. Removing DNA methylation through genetic disruption of DNA methyltransferases (DKO cells) leads to loss of MBD2 occupancy and to a decrease in the frequency of long-range contacts, such that these now more resemble those in undifferentiated Tera-2 cells. Our findings reveal unexpected similarities in higher order chromatin conformation between stem/precursor cells and adult cancers. We also provide novel insight that PcG-occupied and H3K27me3-enriched regions can form chromatin loops and physically interact in cis around a single gene in mammalian cells. The loops associate with a poised, low transcription state in EC cells and, with the addition of DNA methylation, completely repressed transcription in adult cancer cells.
Conflict of interest statement
Figures
Comment in
-
Loopy chromatin brings distant DNA to bear on silencing promoter genes.PLoS Biol. 2008 Dec;6(12):e313. doi: 10.1371/journal.pbio.0060313. Epub 2008 Dec 2. PLoS Biol. 2008. PMID: 20076702 Free PMC article. No abstract available.
References
-
- Sparmann A, van Lohuizen M. Polycomb silencers control cell fate, development and cancer. Nat Rev Cancer. 2006;6:846–856. - PubMed
-
- Ringrose L. Polycomb comes of age: genome-wide profiling of target sites. Curr Opin Cell Biol. 2007;19:290–297. - PubMed
-
- Boyer LA, Plath K, Zeitlinger J, Brambrink T, Medeiros LA, et al. Polycomb complexes repress developmental regulators in murine embryonic stem cells. Nature. 2006;441:349–353. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
