

Structure of a mutant form of proliferating cell nuclear antigen that blocks translesion DNA synthesis
- PMID: 19053247
- PMCID: PMC2703463
- DOI: 10.1021/bi8017762
Structure of a mutant form of proliferating cell nuclear antigen that blocks translesion DNA synthesis
Abstract
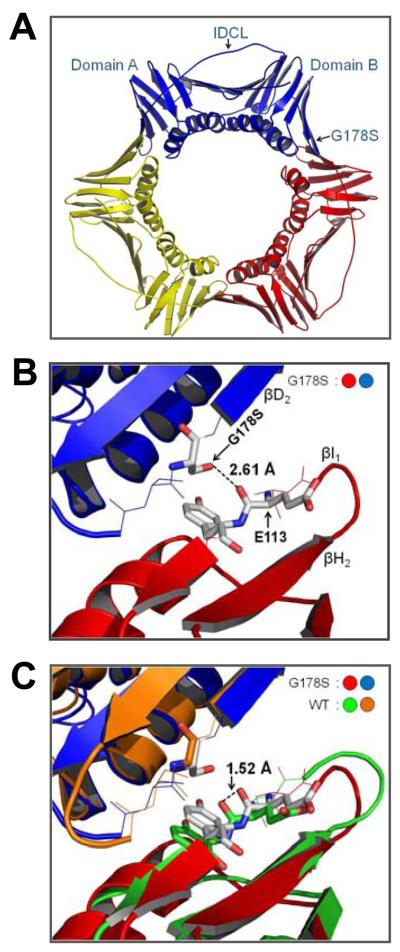
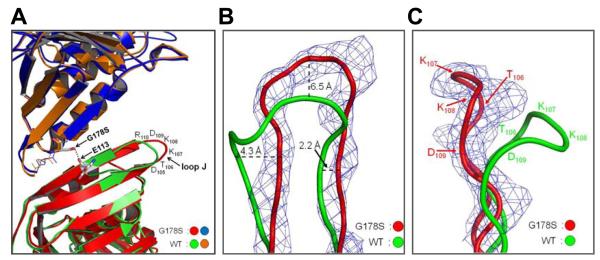
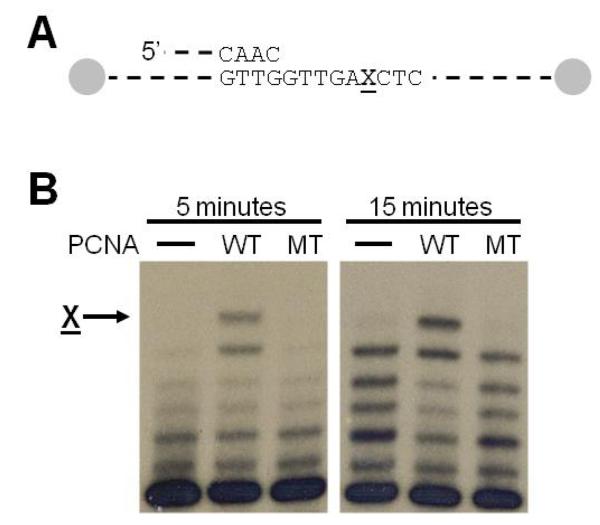
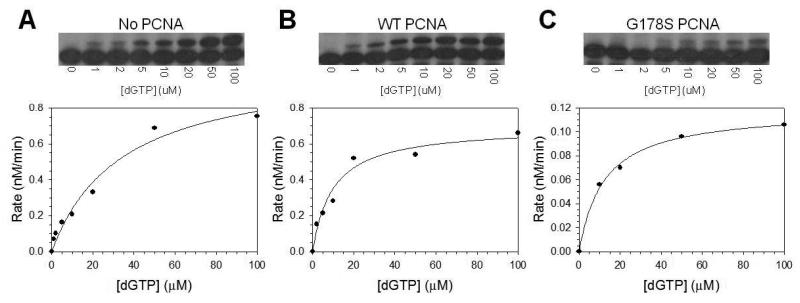
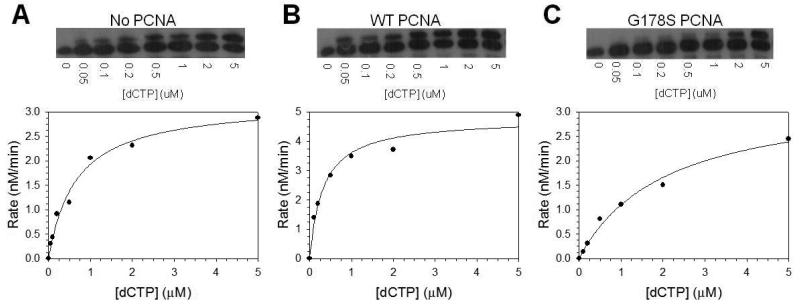
Proliferating cell nuclear antigen (PCNA) is a homotrimeric protein that functions as a sliding clamp during DNA replication. Several mutant forms of PCNA that block translesion DNA synthesis have been identified in genetic studies in yeast. One such mutant protein (encoded by the rev6-1 allele) is a glycine to serine substitution at residue 178, located at the subunit interface of PCNA. To improve our understanding of how this substitution interferes with translesion synthesis, we have determined the X-ray crystal structure of the PCNA G178S mutant protein. This substitution has little effect on the structure of the domain in which the substitution occurs. Instead, significant, local structural changes are observed in the adjacent subunit. The most notable difference between mutant and wild-type structures is in a single, extended loop (comprising amino acid residues 105-110), which we call loop J. In the mutant protein structure, loop J adopts a very different conformation in which the atoms of the protein backbone have moved by as much as 6.5 A from their positions in the wild-type structure. To improve our understanding of the functional consequences of this structural change, we have examined the ability of this mutant protein to stimulate nucleotide incorporation by DNA polymerase eta (pol eta). Steady state kinetic studies show that while wild-type PCNA stimulates incorporation by pol eta opposite an abasic site, the mutant PCNA protein actually inhibits incorporation opposite this DNA lesion. These results show that the position of loop J in PCNA plays an essential role in facilitating translesion synthesis.
Figures

References
-
- Goodman MF. Error-prone repair DNA polymerases in prokaryotes and eukaryotes. Annu. Rev. Biochem. 2002;71:17–50. - PubMed
-
- Prakash S, Prakash L. Translesion DNA synthesis in eukaryotes: a one- or two-polymerase affair. Genes Dev. 2002;16:1872–1883. - PubMed
-
- Prakash S, Johnson RE, Prakash L. Eukaryotic translesion synthesis DNA polymerases: specificity of structure and function. Annu. Rev. Biochem. 2005;74:317–353. - PubMed
-
- Johnson RE, Prakash S, Prakash L. Efficient bypass of a thymine-thymine dimer by yeast DNA polymerase, Pol eta. Science. 1999;283:1001–1004. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
