

CCAAT enhancer-binding protein alpha is a molecular target of 1,25-dihydroxyvitamin D3 in MCF-7 breast cancer cells
- PMID: 19054766
- PMCID: PMC2631956
- DOI: 10.1074/jbc.M803602200
CCAAT enhancer-binding protein alpha is a molecular target of 1,25-dihydroxyvitamin D3 in MCF-7 breast cancer cells
Erratum in
- J Biol Chem. 2009 Mar 20;284(12):8208. Weider, Robert [corrected to Wieder, Robert]
Abstract
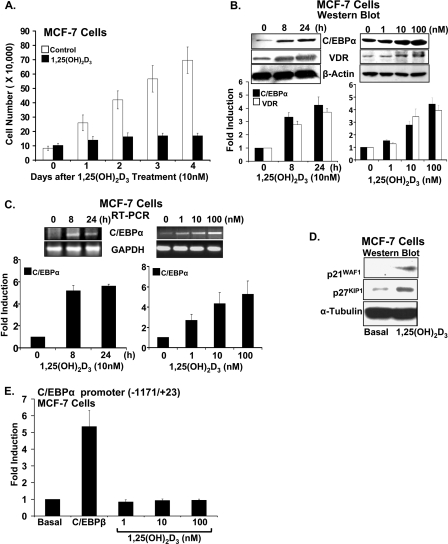
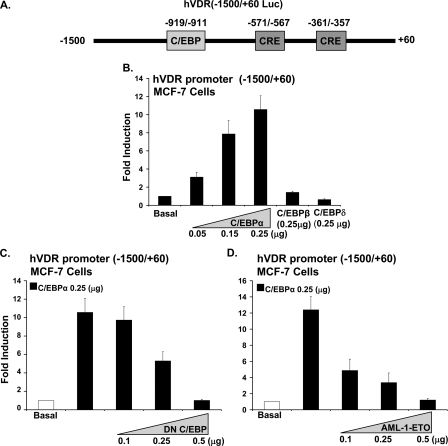
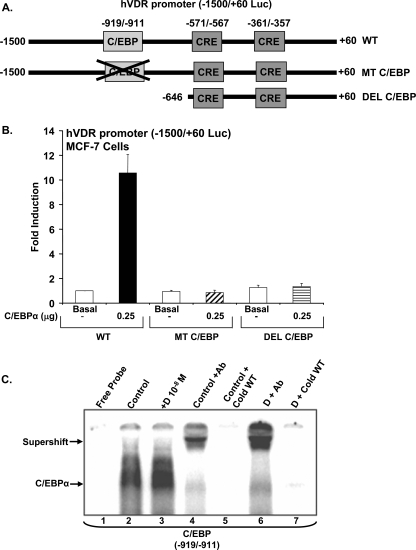
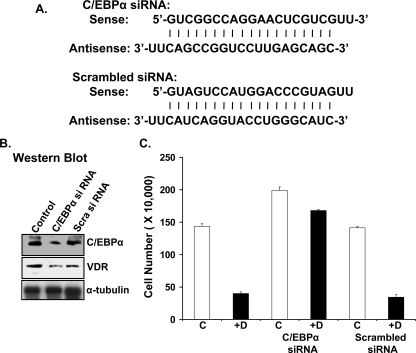
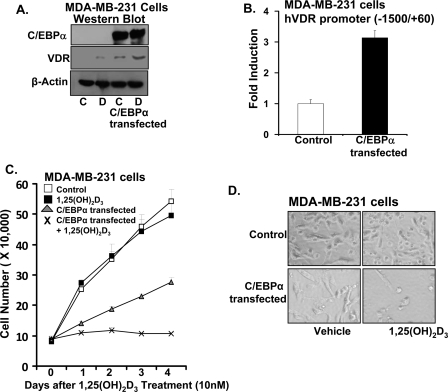
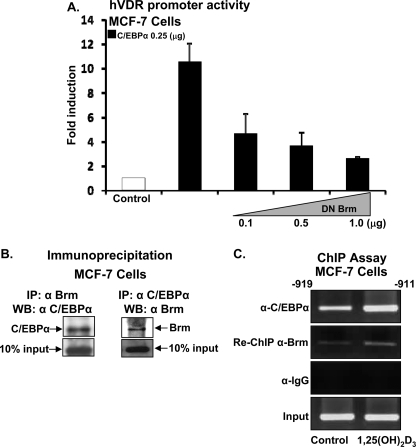
Numerous studies have shown that the active form of vitamin D, 1,25(OH)(2)D(3), can exert growth inhibitory effects on human breast cancer cells and mammary tumor growth. However, the molecular mechanisms remain to be fully delineated. This study demonstrates for the first time that CCAAT enhancer-binding protein alpha (C/EBPalpha), a member of the C/EBP family of transcription factors, is induced by 1,25(OH)(2)D(3) and is a potent enhancer of VDR transcription in MCF-7 breast cancer cells. 1,25(OH)(2)D(3) was found to induce C/EBPalpha as well as VDR expression in MCF-7 cells. C/EBPalpha was not detected in MDA-MB-231 cells that are poorly responsive to 1,25(OH)(2)D(3). Antiproliferative effects of 1,25(OH)(2)D(3) and induction of VDR were observed in MDA-MB-231 cells transfected with C/EBPalpha, and knockdown of C/EBPalpha suppressed VDR and antiproliferative effects of 1,25(OH)(2)D(3) in MCF-7 cells. Transfection of C/EBPalpha in MCF-7 cells resulted in a dose-dependent enhancement of hVDR transcription. Our studies show that C/EBPalpha can bind to Brahma (Brm), an ATPase that is a component of the SWI/SNF complex, and cooperate with Brm in the regulation of hVDR transcription in MCF-7 cells. Because the levels of VDR in MCF-7 breast cancer cells correlate with the antiproliferative effects of 1,25(OH)(2)D(3) and because C/EBPalpha has been suggested as a potential tumor suppressor in breast cancer, these findings provide important mechanisms whereby 1,25(OH)(2)D(3) may act to inhibit growth of breast cancer cells. These findings also identify C/EBPalpha as a 1,25(OH)(2)D(3) target in breast cancer cells and provide evidence for C/EBPalpha as a candidate for breast cancer treatment.
Figures
References
-
- Christakos, S., Dhawan, P., Liu, Y., Peng, X., and Porta, A. (2003) J. Cell Biochem. 88 695-705 - PubMed
-
- Jacobson, E. A., James, K. A., Newmark, H. L., and Carroll, K. K. (1989) Cancer Res. 49 6300-6303 - PubMed
-
- Colston, K. W., Chander, S. K., Mackay, A. G., and Coombes, R. C. (1992) Biochem. Pharmacol. 44 693-702 - PubMed
-
- Colston, K. W., Mackay, A. G., James, S. Y., Binderup, L., Chander, S., and Coombes, R. C. (1992) Biochem. Pharmacol. 44 2273-2280 - PubMed
-
- Anzano, M. A., Smith, J. M., Uskokovic, M. R., Peer, C. W., Mullen, L. T., Letterio, J. J., Welsh, M. C., Shrader, M. W., Logsdon, D. L., and Driver, C. L., Brown, C. C., Roberts A. B., and Sporn, M. B. (1994) Cancer Res. 54 1653-1656 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
