

Modular control of endothelial sheet migration
- PMID: 19056882
- PMCID: PMC2600767
- DOI: 10.1101/gad.1725808
Modular control of endothelial sheet migration
Abstract
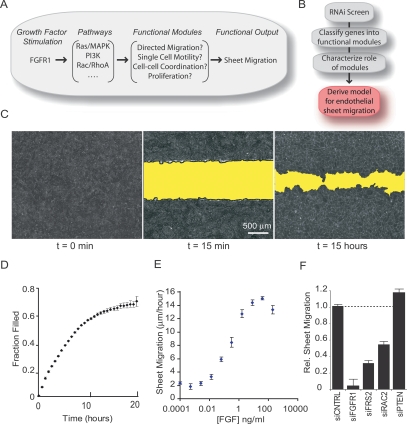
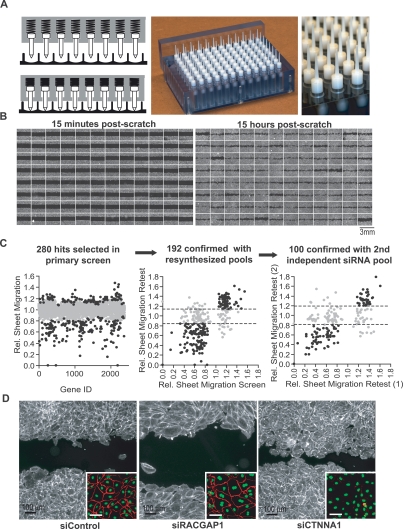
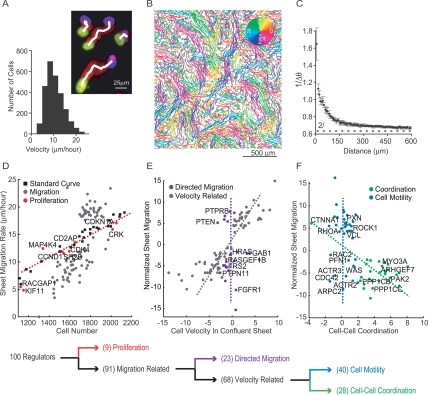
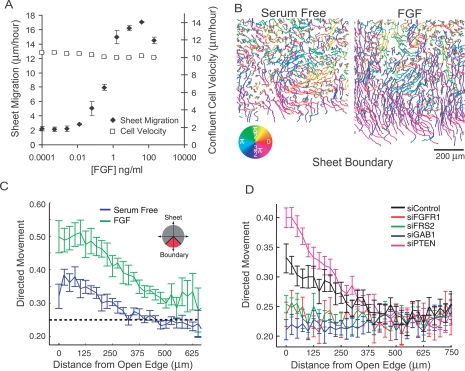
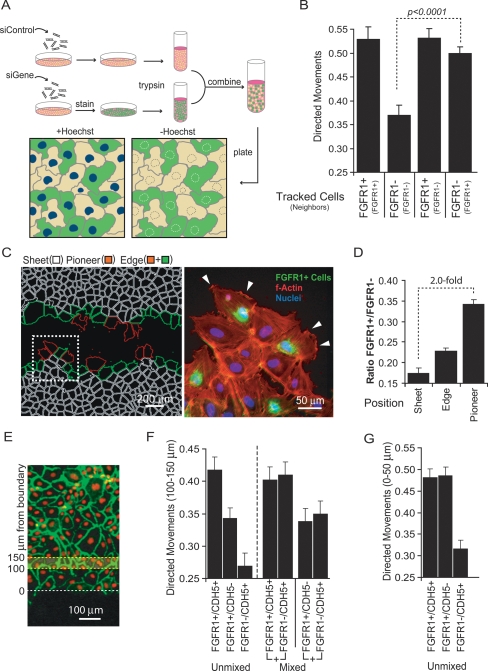
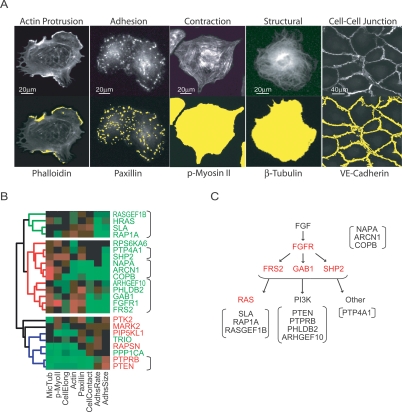
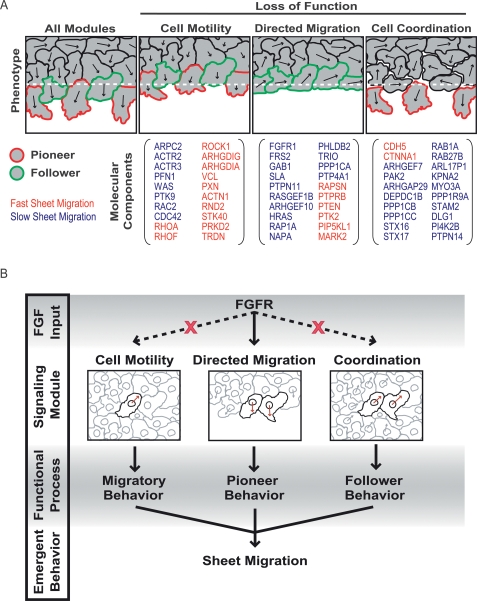
Growth factor-induced migration of endothelial cell monolayers enables embryonic development, wound healing, and angiogenesis. Although collective migration is widespread and therapeutically relevant, the underlying mechanism by which cell monolayers respond to growth factor, sense directional signals, induce motility, and coordinate individual cell movements is only partially understood. Here we used RNAi to identify 100 regulatory proteins that enhance or suppress endothelial sheet migration into cell-free space. We measured multiple live-cell migration parameters for all siRNA perturbations and found that each targeted protein primarily regulates one of four functional outputs: cell motility, directed migration, cell-cell coordination, or cell density. We demonstrate that cell motility regulators drive random, growth factor-independent motility in the presence or absence of open space. In contrast, directed migration regulators selectively transduce growth factor signals to direct cells along the monolayer boundary toward open space. Lastly, we found that regulators of cell-cell coordination are growth factor-independent and reorient randomly migrating cells inside the sheet when boundary cells begin to migrate. Thus, cells transition from random to collective migration through a modular control system, whereby growth factor signals convert boundary cells into pioneers, while cells inside the monolayer reorient and follow pioneers through growth factor-independent migration and cell-cell coordination.
Figures
References
-
- Bikfalvi A., Klein S., Pintucci G., Rifkin D.B. Biological roles of fibroblast growth factor-2. Endocr. Rev. 1997;18:26–45. - PubMed
-
- Bindschadler M., McGrath J.L. Sheet migration by wounded monolayers as an emergent property of single-cell dynamics. J. Cell Sci. 2007;120:876–884. - PubMed
-
- Carmeliet P. Angiogenesis in health and disease. Nat. Med. 2003;9:653–660. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources