

UNC-51/ATG1 kinase regulates axonal transport by mediating motor-cargo assembly
- PMID: 19056884
- PMCID: PMC2600757
- DOI: 10.1101/gad.1734608
UNC-51/ATG1 kinase regulates axonal transport by mediating motor-cargo assembly
Abstract
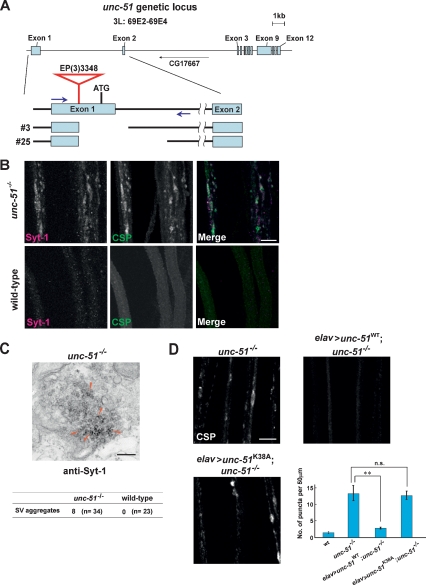
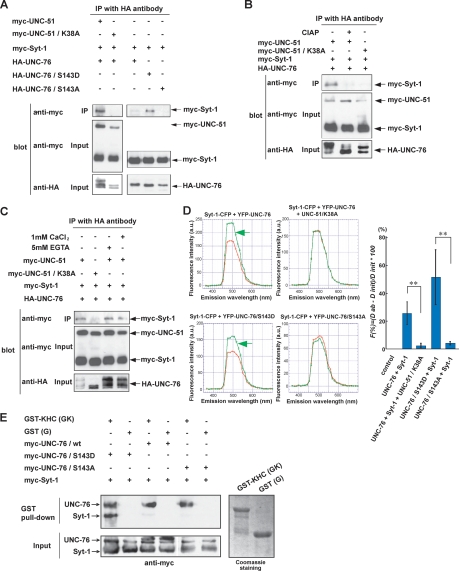
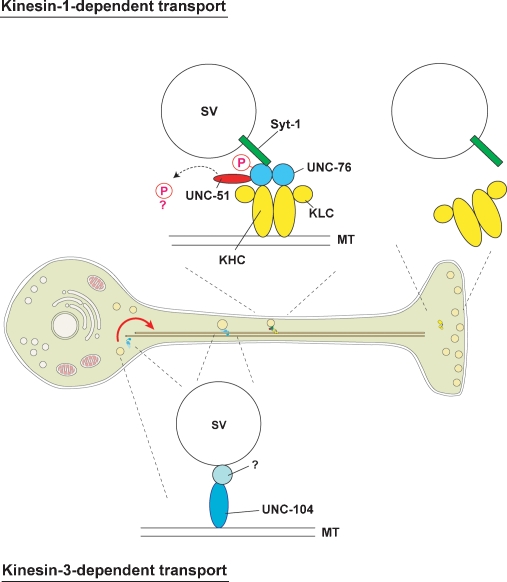
Axonal transport mediated by microtubule-dependent motors is vital for neuronal function and viability. Selective sets of cargoes, including macromolecules and organelles, are transported long range along axons to specific destinations. Despite intensive studies focusing on the motor machinery, the regulatory mechanisms that control motor-cargo assembly are not well understood. Here we show that UNC-51/ATG1 kinase regulates the interaction between synaptic vesicles and motor complexes during transport in Drosophila. UNC-51 binds UNC-76, a kinesin heavy chain (KHC) adaptor protein. Loss of unc-51 or unc-76 leads to severe axonal transport defects in which synaptic vesicles are segregated from the motor complexes and accumulate along axons. Genetic studies show that unc-51 and unc-76 functionally interact in vivo to regulate axonal transport. UNC-51 phosphorylates UNC-76 on Ser(143), and the phosphorylated UNC-76 binds Synaptotagmin-1, a synaptic vesicle protein, suggesting that motor-cargo interactions are regulated in a phosphorylation-dependent manner. In addition, defective axonal transport in unc-76 mutants is rescued by a phospho-mimetic UNC-76, but not a phospho-defective UNC-76, demonstrating the essential role of UNC-76 Ser(143) phosphorylation in axonal transport. Thus, our data provide insight into axonal transport regulation that depends on the phosphorylation of adaptor proteins.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases