

Chromatin structure analyses identify miRNA promoters
- PMID: 19056895
- PMCID: PMC2593607
- DOI: 10.1101/gad.1706508
Chromatin structure analyses identify miRNA promoters
Abstract
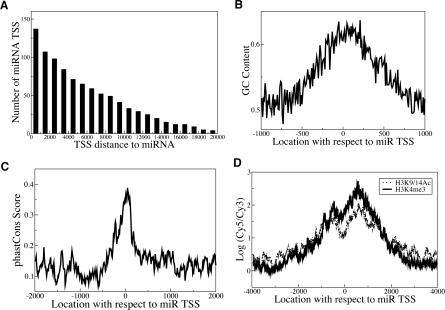
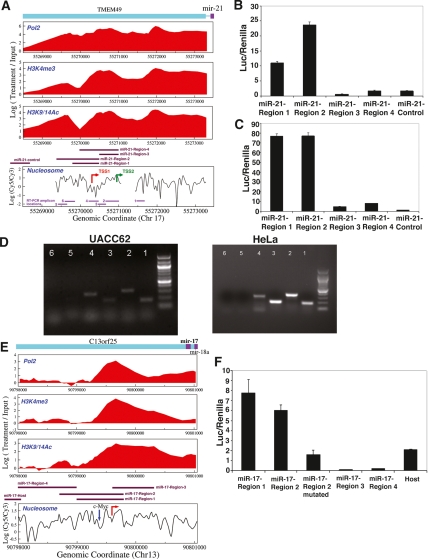
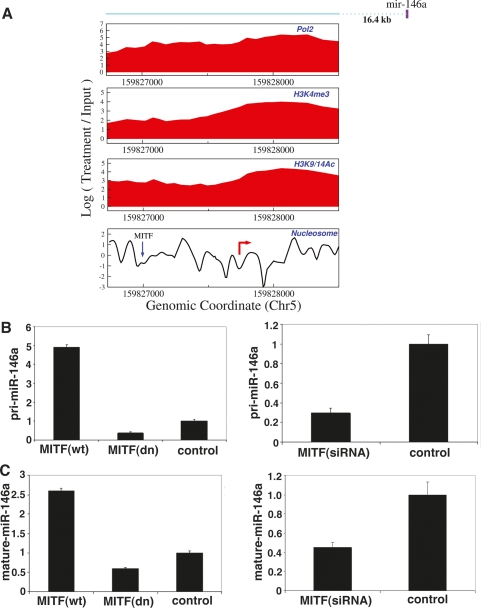
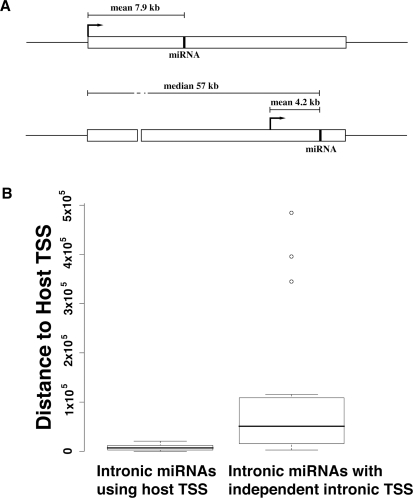
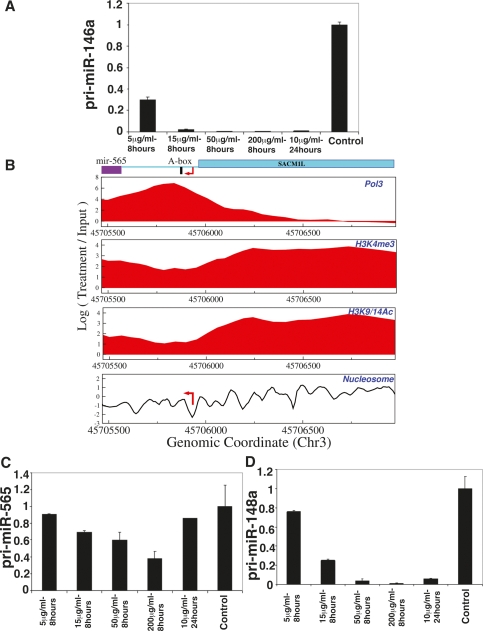
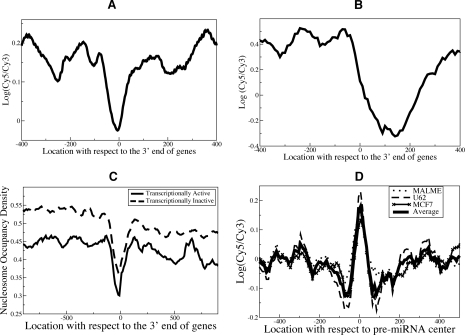
Although microRNAs (miRNAs) are key regulators of gene expression in normal human physiology and disease, transcriptional regulation of miRNAs is poorly understood, because most miRNA promoters have not yet been characterized. We identified the proximal promoters of 175 human miRNAs by combining nucleosome mapping with chromatin signatures for promoters. We observe that one-third of intronic miRNAs have transcription initiation regions independent from their host promoters and present a list of RNA polymerase II- and III-occupied miRNAs. Nucleosome mapping and linker sequence analyses in miRNA promoters permitted accurate prediction of transcription factors regulating miRNA expression, thus identifying nine miRNAs regulated by the MITF transcription factor/oncoprotein in melanoma cells. Furthermore, DNA sequences encoding mature miRNAs were found to be preferentially occupied by positioned-nucleosomes, and the 3' end sites of known genes exhibited nucleosome depletion. The high-throughput identification of miRNA promoter and enhancer regulatory elements sheds light on evolution of miRNA transcription and permits rapid identification of transcriptional networks of miRNAs.
Figures
References
-
- Alvarez-Garcia I., Miska E.A. MicroRNA functions in animal development and human disease. Development. 2005;132:4653–4662. - PubMed
-
- Ambros V. The functions of animal microRNAs. Nature. 2004;431:350–355. - PubMed
-
- Anderson J.D., Widom J. Sequence and position-dependence of the equilibrium accessibility of nucleosomal DNA target sites. J. Mol. Biol. 2000;296:979–987. - PubMed
-
- Aravin A.A., Lagos-Quintana M., Yalcin A., Zavolan M., Marks D., Snyder B., Gaasterland T., Meyer J., Tuschl T. The small RNA profile during Drosophila melanogaster development. Dev. Cell. 2003;5:337–350. - PubMed
-
- Barski A., Cuddapah S., Cui K., Roh T.Y., Schones D.E., Wang Z., Wei G., Chepelev I., Zhao K. High-resolution profiling of histone methylations in the human genome. Cell. 2007;129:823–837. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases