

Targeting the X chromosome during spermatogenesis induces Y chromosome transmission ratio distortion and early dominant embryo lethality in Anopheles gambiae
- PMID: 19057670
- PMCID: PMC2585807
- DOI: 10.1371/journal.pgen.1000291
Targeting the X chromosome during spermatogenesis induces Y chromosome transmission ratio distortion and early dominant embryo lethality in Anopheles gambiae
Abstract
We have exploited the high selectivity of the homing endonuclease I-PpoI for the X-linked Anopheles gambiae 28S ribosomal genes to selectively target X chromosome carrying spermatozoa. Our data demonstrated that in heterozygous males, the expression of I-PpoI in the testes induced a strong bias toward Y chromosome-carrying spermatozoa. Notably, these male mosquitoes also induced complete early dominant embryo lethality in crosses with wild-type females. Morphological and molecular data indicated that all spermatozoa, irrespectively of the inheritance of the transgene, carried a substantial amount of I-PpoI protein that could attack the maternally inherited chromosome X of the embryo. Besides the obvious implications for implementing vector control measures, our data demonstrated the feasibility of generating synthetic sex distorters and revealed the intriguing possibility of manipulating maternally inherited genes using wild-type sperm cells carrying engineered endonucleases.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
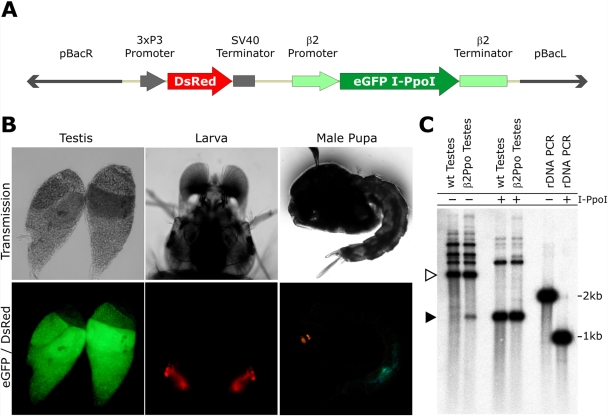

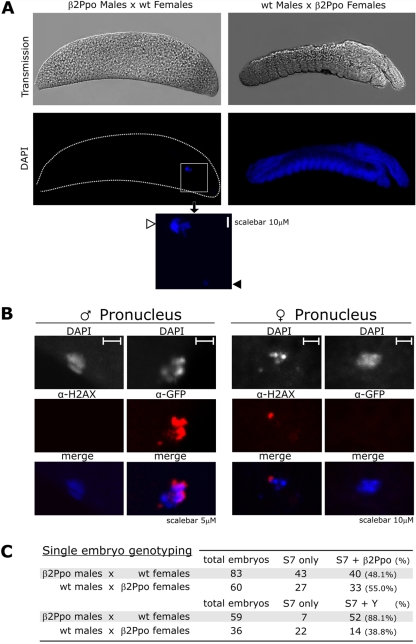
References
-
- World Health Organisation. Geneva: WHO; 2005. World Malaria Report 2005: Roll Back Malaria.
-
- Curtis CF, Graves PM. Methods for replacement of malaria vector populations. J Trop Med Hyg. 1988;91:43–48. - PubMed
-
- Carlson J, Olson K, Higgs S, Beaty B. Molecular genetic manipulation of mosquito vectors. Annu Rev Entomol. 1995;40:359–388. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
