

Topoisomerase II: a fitted mechanism for the chromatin landscape
- PMID: 19059997
- PMCID: PMC2647320
- DOI: 10.1093/nar/gkn994
Topoisomerase II: a fitted mechanism for the chromatin landscape
Abstract
The mechanism by which type-2A topoisomerases transport one DNA duplex through a transient double-strand break produced in another exhibits fascinating traits. One of them is the fine coupling between inter-domainal movements and ATP usage; another is their preference to transport DNA in particular directions. These capabilities have been inferred from in vitro studies but we ignore their significance inside the cell, where DNA configurations markedly differ from those of DNA in free solution. The eukaryotic type-2A enzyme, topoisomerase II, is the second most abundant chromatin protein after histones and its biological roles include the decatenation of newly replicated DNA and the relaxation of polymerase-driven supercoils. Yet, topoisomerase II is also implicated in other cellular processes such as chromatin folding and gene expression, in which the topological transformations catalysed by the enzyme are uncertain. Here, some capabilities of topoisomerase II that might be relevant to infer the enzyme performance in the context of chromatin architecture are discussed. Some aspects addressed are the importance of the DNA rejoining step to ensure genome stability, the regulation of the enzyme activity and of its putative structural role, and the selectively of DNA transport in the chromatin milieu.
Figures
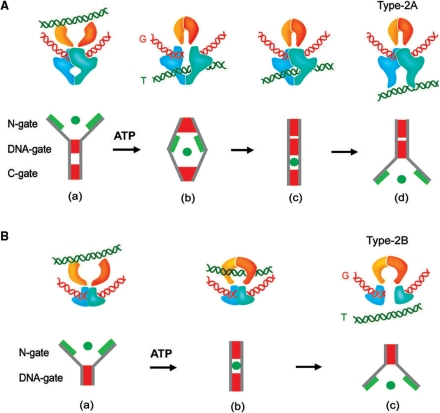



References
-
- Wang JC. Moving one DNA double helix through another by a type II DNA topoisomerase: the story of a simple molecular machine. Q. Rev. Biophys. 1998;31:107–144. - PubMed
-
- Champoux JJ. DNA topoisomerases: structure, function, and mechanism. Annu. Rev. Biochem. 2001;70:369–413. - PubMed
-
- Corbett KD, Berger JM. Structure, molecular mechanisms, and evolutionary relationships in DNA topoisomerases. Annu. Rev. Biophys. Biomol. Struct. 2004;33:95–118. - PubMed
-
- Schoeffler AJ, Berger JM. DNA topoisomerases: harnessing and constraining energy to govern chromosome topology. Q. Rev. Biophys. 2008;41:41–101. - PubMed
