

Interactions of human O(6)-alkylguanine-DNA alkyltransferase (AGT) with short double-stranded DNAs
- PMID: 19061338
- PMCID: PMC2801566
- DOI: 10.1021/bi801666c
Interactions of human O(6)-alkylguanine-DNA alkyltransferase (AGT) with short double-stranded DNAs
Abstract
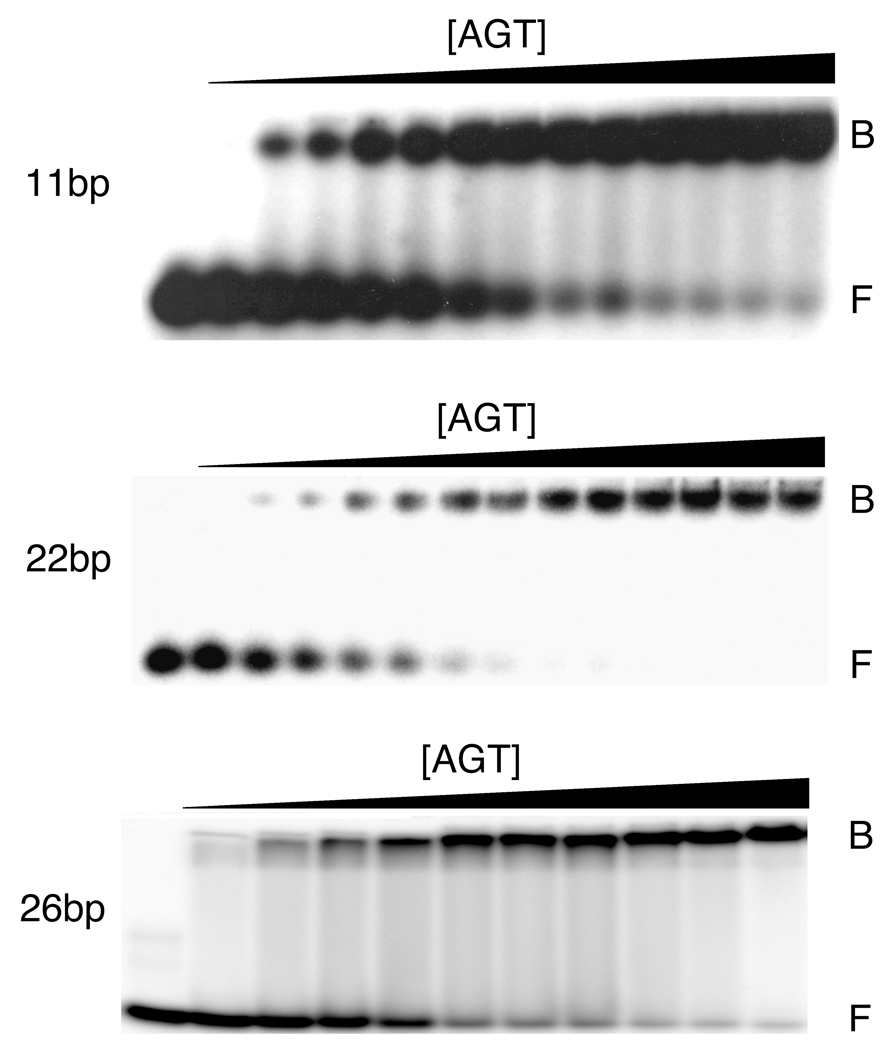
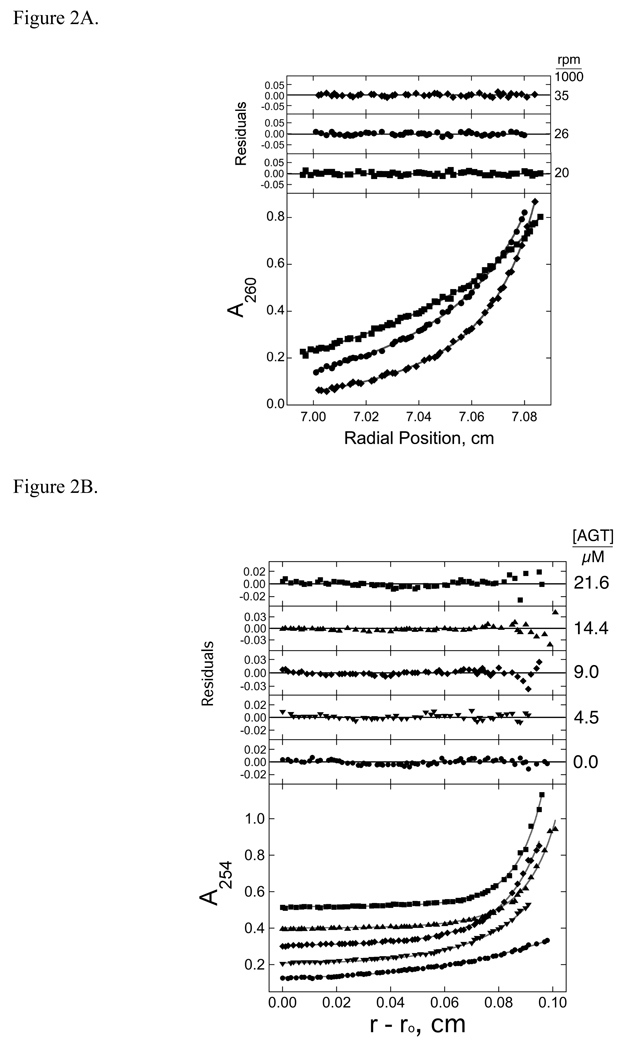
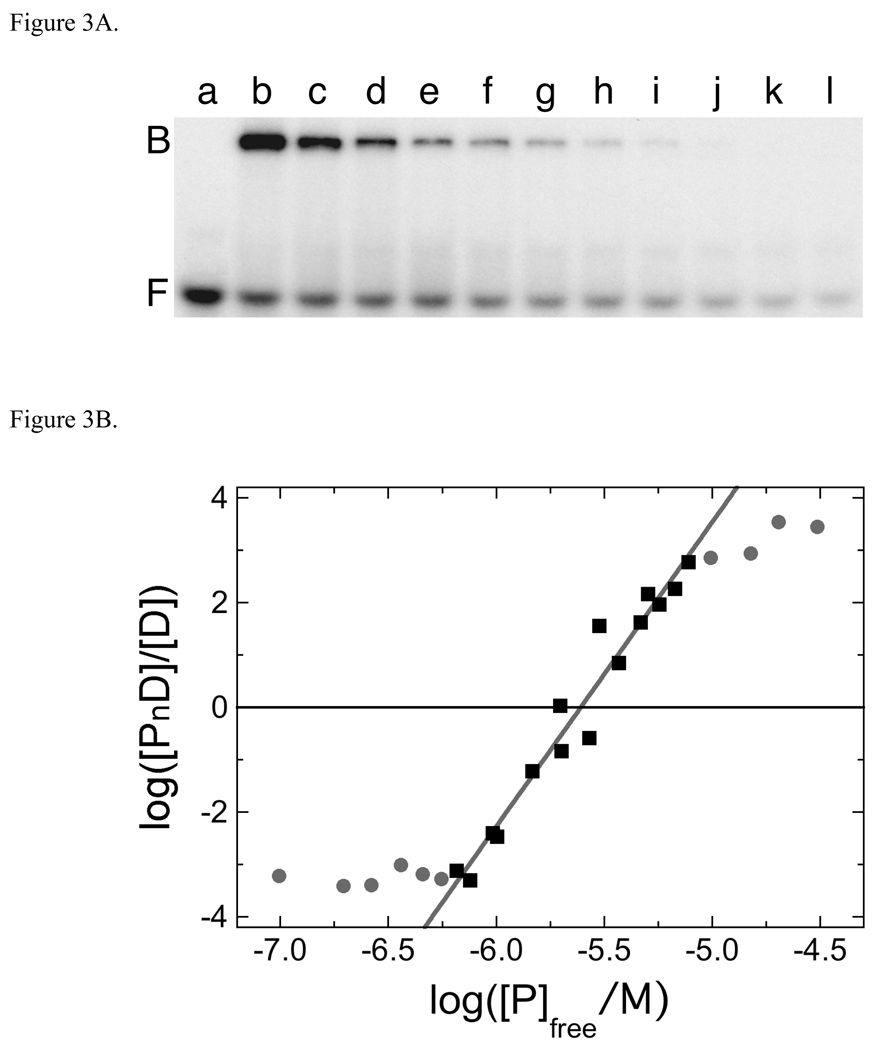
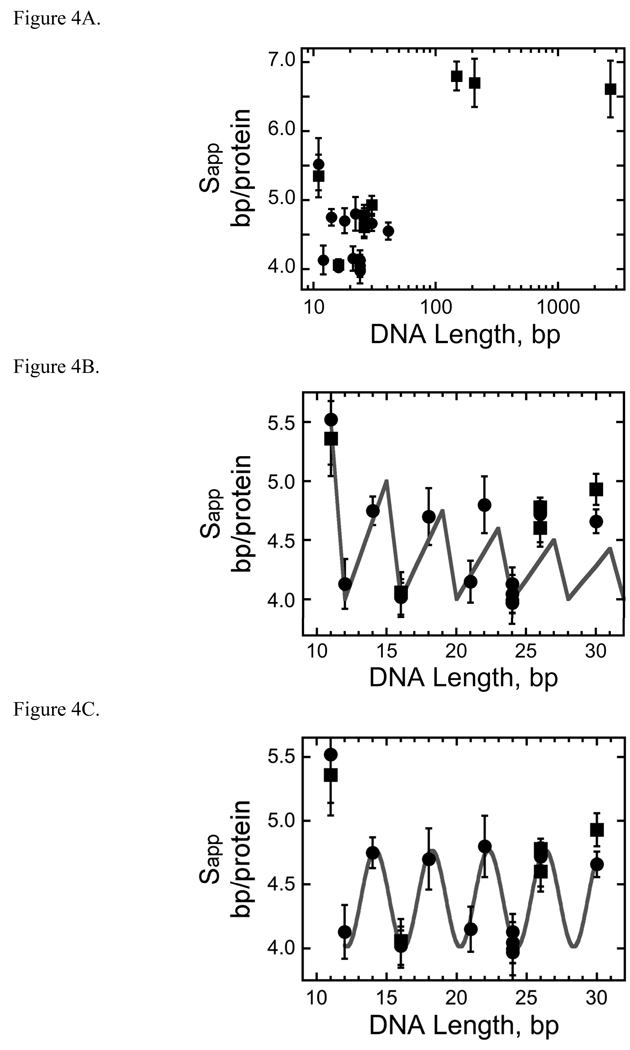
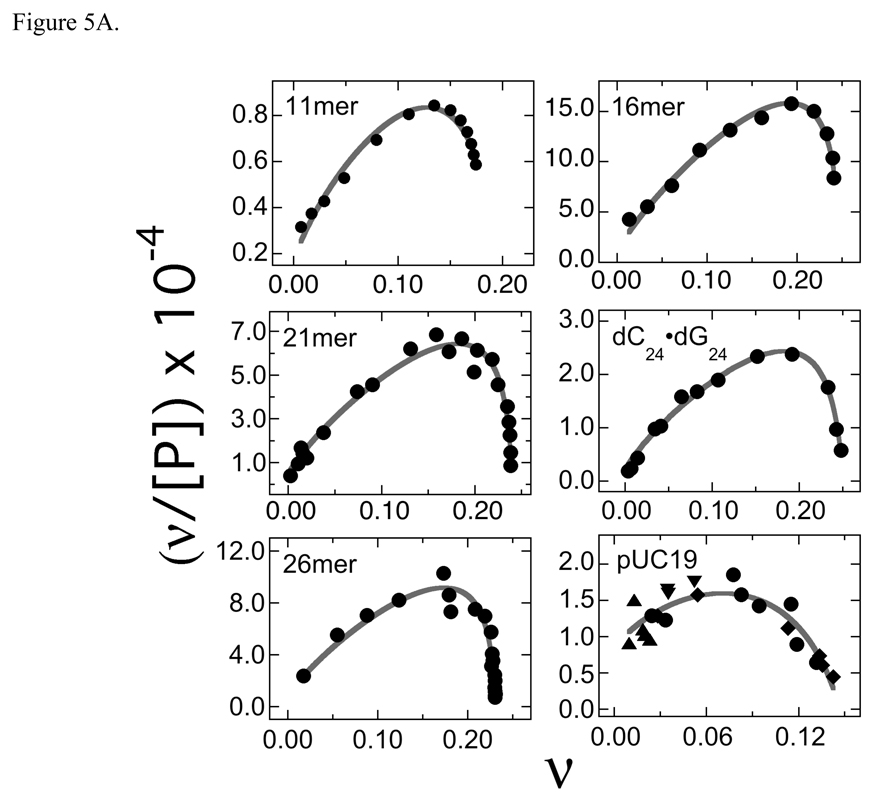
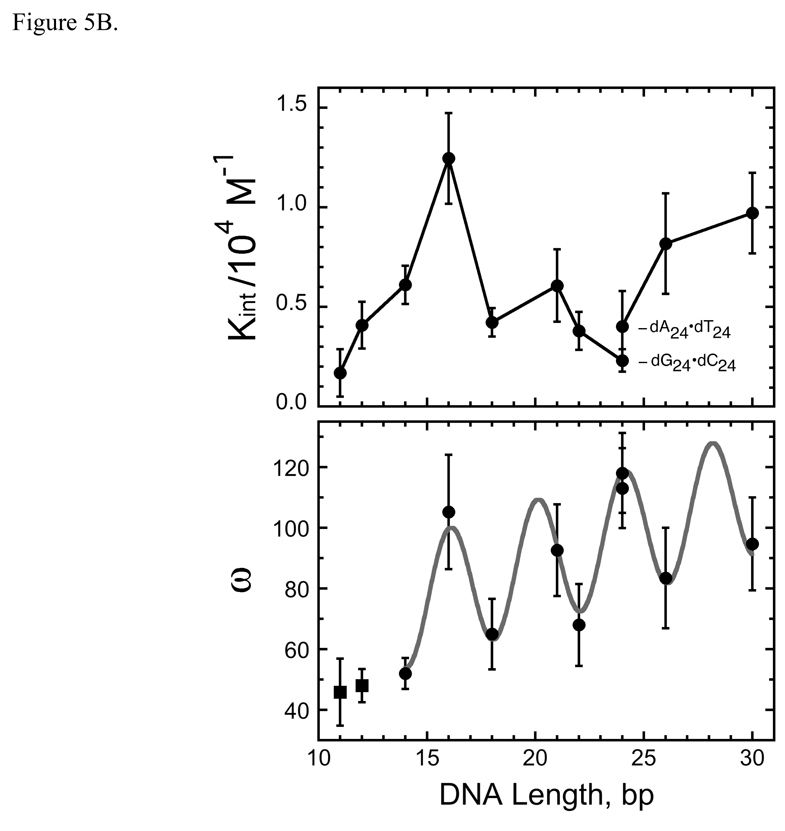
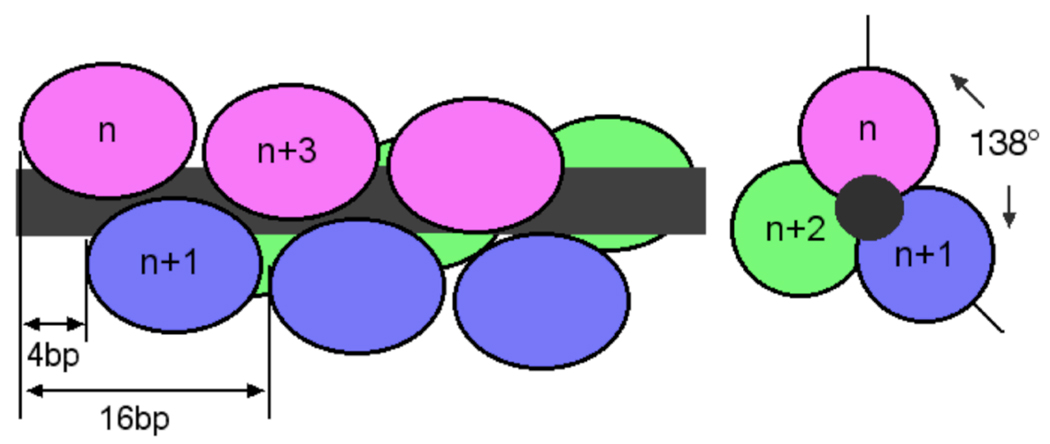
O(6)-alkylguanine-DNA alkyltransferase (AGT) is a ubiquitous enzyme with an amino acid sequence that is conserved in Eubacteria, Archaea, and Eukarya. It repairs O(6)-alkylguanine and O(4)-alkylthymine adducts in single-stranded and duplex DNAs. In performing these functions, AGT must partition between adduct-containing sites and the large excess of adduct-free DNA distributed throughout the genome. Here, we characterize the binding of human AGT to linear double-stranded, adduct-free DNAs ranging in length from 11 bp to 2686 bp. Moderately cooperative binding (22.6 +/- 3.7 < or = omega < or = 145.0 +/- 37.0) results in an all-or-nothing association pattern on short templates. The apparent binding site size S(app) (mean = 4.39 +/- 0.02 bp) oscillates with increasing template length. Oscillations in cooperativity factor omega have the same frequency but are of opposite phase to S(app), with the result that the most stable protein-protein and protein-DNA interactions occur at the highest packing densities. The oscillation period (4.05 +/- 0.02 bp/protein) is nearly identical to the occluded binding site size obtained at the highest measured binding density (4 bp/protein) and is significantly smaller than the contour length ( approximately 8 bp) occupied in crystalline complexes. A model in which protein molecules overlap along the DNA contour is proposed to account for these features. High AGT densities resulting from cooperative binding may allow efficient search for lesions in the context of chromatin remodeling and DNA replication.
Figures
References
-
- Loveless A. Nature (London) Vol. 223. 1969. Possible relevance of O6 alkylation of dexoyguanosine to he mutagenicity and carcinogenicity of nitrosamines and nitrosamides; pp. 206–207. - PubMed
-
- Pegg AE. Methylation of the O6 position of guanine in DNA is the most likely initiating event in carcinogenesis by methylating agents. Cancer Invest. 1984;2:223–231. - PubMed
-
- Pegg AE, Dolan ME, Moschel RC. Structure, function and inhibition of O6-alkylguanine-DNA alkyltransferase. Prog. Nucl. Acid Res. and Mol. Biol. 1995;51:167–223. - PubMed
-
- Pegg AE. Mammalian O6-alkylguanine-DNA alkyltransferase: regulation and importance in response to alkylating carcinogens and therapeutic agents. Cancer Res. 1990;50:6119–6129. - PubMed
-
- Margison GP, Santibáñez-Koref MF. O6-alkylguanine-DNA alkyltransferase: role in carcinogenesis and chemotherapy. BioEssays. 2002;24:255–266. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
