

Psoralen-induced DNA interstrand cross-links block transcription and induce p53 in an ataxia-telangiectasia and rad3-related-dependent manner
- PMID: 19064630
- PMCID: PMC2684911
- DOI: 10.1124/mol.108.051698
Psoralen-induced DNA interstrand cross-links block transcription and induce p53 in an ataxia-telangiectasia and rad3-related-dependent manner
Abstract
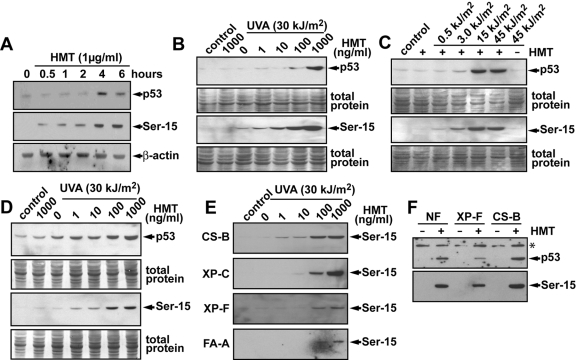
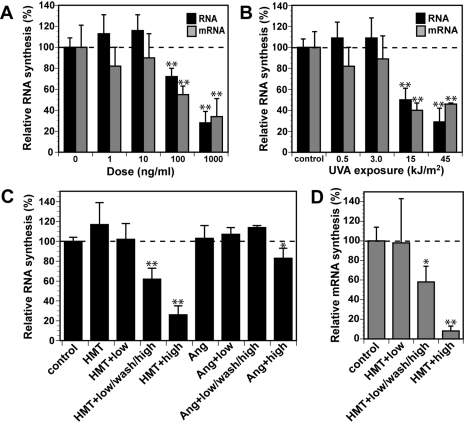
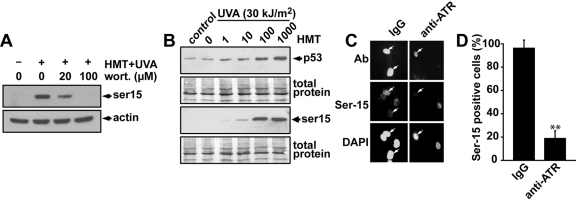
Psoralen plus UVA light (PUVA) is commonly used to treat psoriasis, a common skin disorder associated with rapid proliferation of cells. PUVA exerts its antiproliferative activity through formation of DNA monoadducts and interstrand cross-links (ICLs). However, this treatment may lead to skin malignancies as a direct result of inducing carcinogenic DNA damage. Inactivation of the p53 tumor suppressor gene is an important event in the development of skin cancer. p53 is rapidly phosphorylated and stabilized in response to DNA damage, and the induction of apoptosis by p53 is an important mechanism by which p53 exerts its tumor-suppressive activity. To better understand the mechanism by which PUVA treatment induces p53, we exposed human skin fibroblasts with PUVA under conditions that differentially produce monoadducts and ICLs and found that psoralen-induced ICLs induced phosphorylation of the Ser-15 site of p53 and apoptosis much more effectively than psoralen-induced monoadducts. The induction of p53 phosphorylation by psoralen ICLs did not require factors believed to be involved in the repair of psoralen ICLs [xeroderma pigmentosum (XP)-A, XP-C, XP-F, Cockayne's syndrome-B, Fanconi anemia] but did require the ataxia-telangiectasia and Rad3-related but not the ataxia-telangiectasia mutated kinase. Psoralen-induced ICLs blocked transcription and replication more efficiently than monoadducts, and induction of p53 and apoptosis correlated with doses causing interference with transcription rather than DNA replication. Our finding that cells underwent apoptosis preferentially during S-phase suggests that the combined blockade of transcription and DNA replication by psoralen ICLs during S-phase elicits a strong apoptotic response.
Figures


References
-
- Bessho T (2003) Induction of DNA replication-mediated double strand breaks by psoralen DNA interstrand cross-links. J Biol Chem 278 5250-5254. - PubMed
-
- Bode AM and Dong Z (2004) Post-translational modification of p53 in tumorigenesis. Nat Rev Cancer 4 793-805. - PubMed
-
- Brash DE, Wikonkal NM, Remenyik E, van der Horst GT, Friedberg EC, Cheo DL, van Steeg H, Westerman A, and van Kranen HJ (2001) The DNA damage signal for Mdm2 regulation, Trp53 induction, and sunburn cell formation in vivo originates from actively transcribed genes. J Invest Dermatol 117 1234-1240. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
