

Leishmania donovani ornithine decarboxylase is indispensable for parasite survival in the mammalian host
- PMID: 19064633
- PMCID: PMC2632046
- DOI: 10.1128/IAI.01236-08
Leishmania donovani ornithine decarboxylase is indispensable for parasite survival in the mammalian host
Abstract
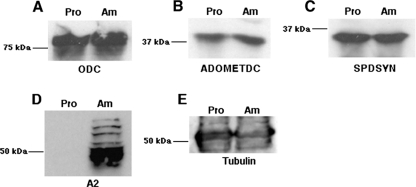
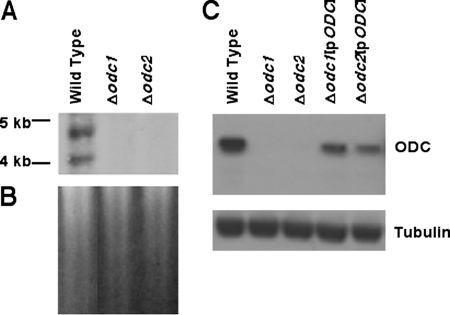
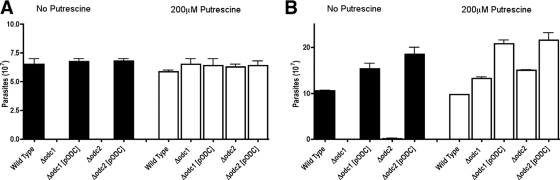
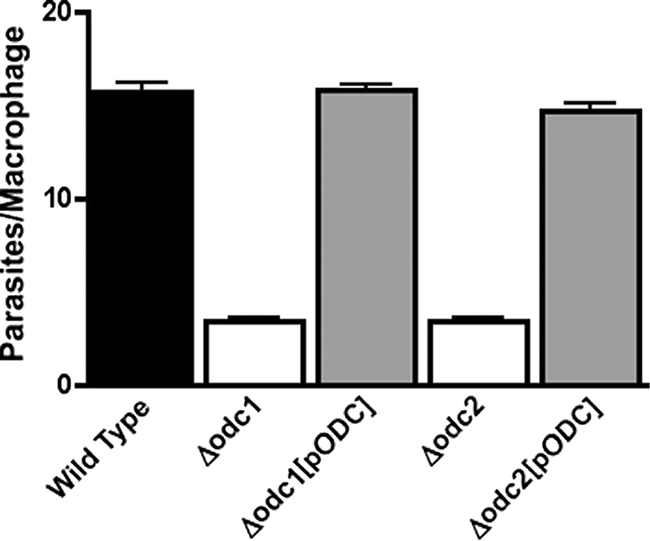
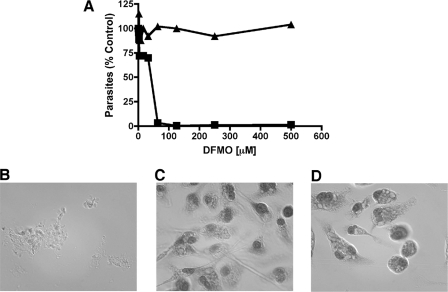
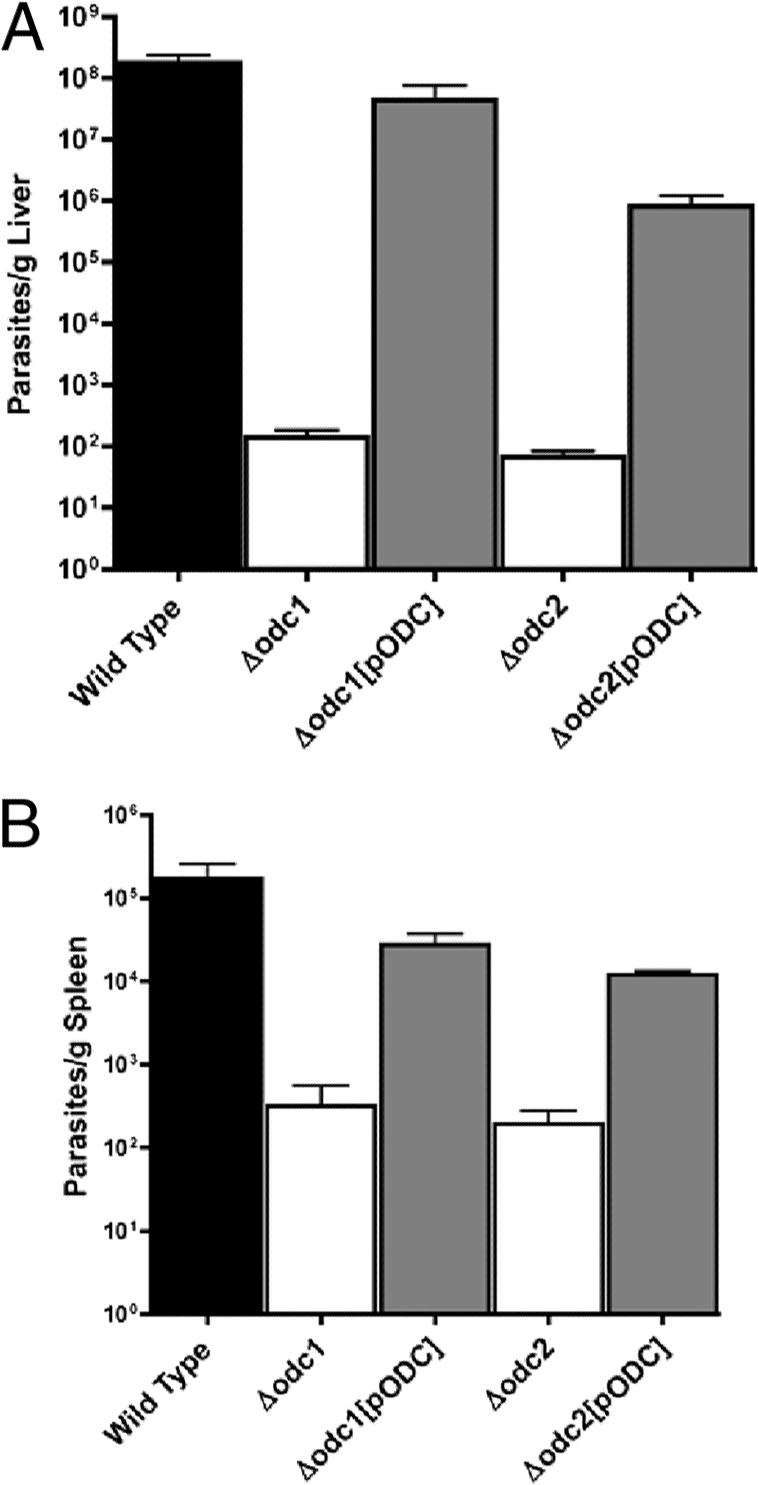
Mutations within the polyamine biosynthetic pathway of Leishmania donovani, the etiological agent of visceral leishmaniasis, confer polyamine auxotrophy to the insect vector or promastigote form of the parasite. However, whether the infectious or amastigote form of the parasite requires an intact polyamine pathway has remained an open question. To address this issue, conditionally lethal Deltaodc mutants lacking ornithine decarboxylase (ODC), the rate-limiting enzyme in polyamine biosynthesis, were created by double targeted gene replacement within a virulent strain of L. donovani. ODC-deficient promastigotes and axenic amastigotes were auxotrophic for polyamines and capable of robust growth only when exogenous putrescine was supplied in the culture medium, confirming that polyamine biosynthesis is an essential nutritional pathway for L. donovani promastigotes. To assess whether the Deltaodc lesion also affected the ability of amastigotes to sustain a robust infection, macrophage and mouse infectivity experiments were performed. Parasite loads in murine macrophages infected with each of two independent Deltaodc knockout lines were decreased approximately 80% compared to their wild-type counterpart. Furthermore, alpha-difluoromethylornithine, a suicide inhibitor of ODC, inhibited growth of wild-type L. donovani amastigotes and effectively cured macrophages of parasites, thereby preventing host cell destruction. Strikingly, however, parasitemias of both Deltaodc null mutants were reduced by 6 and 3 orders of magnitude, respectively, in livers and spleens of BALB/c mice. The compromised infectivity phenotypes of the Deltaodc knockouts in both macrophages and mice were rescued by episomal complementation of the genetic lesion. These genetic and pharmacological studies strongly implicate ODC as an essential cellular determinant that is necessary for the viability and growth of both L. donovani promastigotes and amastigotes and intimate that pharmacological inhibition of ODC is a promising therapeutic paradigm for the treatment of visceral and perhaps other forms of leishmaniasis.
Figures
References
-
- Assaraf, Y. G., J. Golenser, D. T. Spira, G. Messer, and U. Bachrach. 1987. Cytostatic effect of DL-alpha-difluoromethylornithine against Plasmodium falciparum and its reversal by diamines and spermidine. Parasitol. Res. 73313-318. - PubMed
-
- Bacchi, C. J., and P. P. McCann. 1987. Parasitic protozoa and polyamines, p. 317-344. In P. P. McCann, A. E. Pegg, and A. Sjoerdsma (ed.), Inhibition of polyamine metabolism: biological significance and basis for new therapies. Academic Press, Orlando, FL.
-
- Bacchi, C. J., and N. Yarlett. 1995. Polyamine metabolism. In J. J. Marr and M. Muller (ed.), Biochemistry and molecular biology of parasites, p. 119-131. Academic Press Ltd., London, United Kingdom.
-
- Bachrach, U. 2005. Naturally occurring polyamines: interaction with macromolecules. Curr. Protein Pept. Sci. 6559-566. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
