

Plasma membrane microdomains regulate turnover of transport proteins in yeast
- PMID: 19064668
- PMCID: PMC2600745
- DOI: 10.1083/jcb.200806035
Plasma membrane microdomains regulate turnover of transport proteins in yeast
Abstract
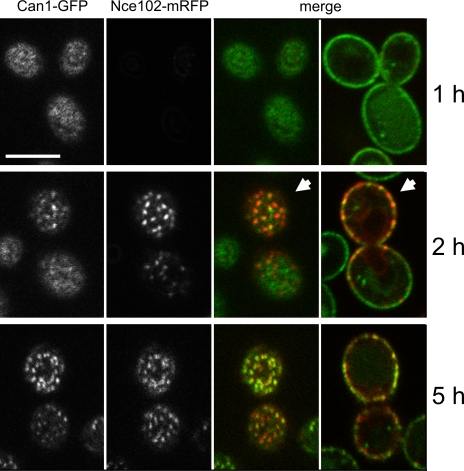
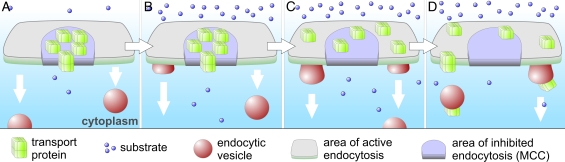
In this study, we investigate whether the stable segregation of proteins and lipids within the yeast plasma membrane serves a particular biological function. We show that 21 proteins cluster within or associate with the ergosterol-rich membrane compartment of Can1 (MCC). However, proteins of the endocytic machinery are excluded from MCC. In a screen, we identified 28 genes affecting MCC appearance and found that genes involved in lipid biosynthesis and vesicle transport are significantly overrepresented. Deletion of Pil1, a component of eisosomes, or of Nce102, an integral membrane protein of MCC, results in the dissipation of all MCC markers. These deletion mutants also show accelerated endocytosis of MCC-resident permeases Can1 and Fur4. Our data suggest that release from MCC makes these proteins accessible to the endocytic machinery. Addition of arginine to wild-type cells leads to a similar redistribution and increased turnover of Can1. Thus, MCC represents a protective area within the plasma membrane to control turnover of transport proteins.
Figures








References
-
- Bagnat, M., and K. Simons. 2002. Lipid rafts in protein sorting and cell polarity in budding yeast Saccharomyces cerevisiae. Biol. Chem. 383:1475–1480. - PubMed
-
- Brown, D.A., and J.K. Rose. 1992. Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell surface. Cell. 68:533–544. - PubMed
-
- Bullock, W.O., J.M. Fernandez, and J.M. Short. 1987. Xl1-Blue: a high efficiency plasmid transforming recA Escherichia coli strain with beta-galactosidase selection. Biotechniques. 5:376–379.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
